# Tak zwane jednostki chronostratygraficzne (skalne), które nie wiedzieć po co przeciwstawia się jednostkom geochronologicznym (czasowym), choć mają te same nazwy i ten sam zakres.
* Do niedawna nazwa nieformalna, w starszej literaturze spotyka się też nazwy prearchaik, azoik, priskoik lub katarchaik. Według jednego z szacunków hadeik zaczął się 4567,17 mlt.
| Rank | Name | Common name | Started (millions of years ago) |
|---|---|---|---|
| Life | 4,200 | ||
| Archaea | 3,700 | ||
| Domain | Eukaryota | Eukaryotes | 2,100 |
| Opimoda | Excludes Plants and their relatives | 1,540 | |
| Amorphea | |||
| Obazoa | Excludes Amoebozoa (Amoebas) | ||
| Opisthokonta | Holozoa + Holomycota (Cristidicoidea and Fungi) | 1,300 | |
| Holozoa | Excludes Holomycota | 1,100 | |
| Filozoa | Choanozoa + Filasterea | ||
| Choanozoa | Choanoflagellates + Animals | 900 | |
| Kingdom | Animalia | Animals | 610 |
| Subkingdom | Eumetazoa | Excludes Porifera (Sponges) | |
| Parahoxozoa | Excludes Ctenophora (Comb Jellies) | ||
| Bilateria | Triploblasts / Worms | 560 | |
| Nephrozoa | |||
| Deuterostomia | Division from Protostomes | ||
| Phylum | Chordata | Chordates (Vertebrates and closely related invertebrates) | 530 |
| Olfactores | Excludes cephalochordates (Lancelets) | ||
| Subphylum | Vertebrata | Fish / Vertebrates | 505 |
| Infraphylum | Gnathostomata | Jawed fish | 460 |
| Teleostomi | Bony fish | 420 | |
| Sarcopterygii | Lobe finned fish | ||
| Superclass | Tetrapoda | Tetrapods (animals with four limbs) | 395 |
| Amniota | Amniotes (fully terrestrial tetrapods whose eggs are "equipped with an amnion") | 340 | |
| Synapsida | Proto-Mammals | 308 | |
| Therapsida | Limbs beneath the body and other mammalian traits | 280 | |
| Class | Mammalia | Mammals | 220 |
| Subclass | Theria | Mammals that give birth to live young (i.e. non-egg-laying) | 160 |
| Infraclass | Eutheria | Placental mammals (i.e. non-marsupials) | 125 |
| Magnorder | Boreoeutheria | Supraprimates, (most) hoofed mammals, (most) carnivorous mammals, cetaceans, and bats | 124–101 |
| Superorder | Euarchontoglires | Supraprimates: primates, colugos, tree shrews, rodents, and rabbits | 100 |
| Grandorder | Euarchonta | Primates, colugos, and tree shrews | 99–80 |
| Mirorder | Primatomorpha | Primates and colugos | 79.6 |
| Order | Primates | Primates / Plesiadapiformes | 66 |
| Suborder | Haplorrhini | "Dry-nosed" (literally, "simple-nosed") primates: tarsiers and monkeys (incl. apes) | 63 |
| Infraorder | Simiiformes | monkeys (incl. apes) | 40 |
| Parvorder | Catarrhini | "Downward-nosed" primates: apes and old-world monkeys | 30 |
| Superfamily | Hominoidea | Apes: great apes and lesser apes (gibbons) | 22–20 |
| Family | Hominidae | Great apes: humans, chimpanzees, gorillas and orangutans—the hominids | 20–15 |
| Subfamily | Homininae | Humans, chimpanzees, and gorillas (the African apes) | 14–12 |
| Tribe | Hominini | Includes both Homo and Pan (chimpanzees), but not Gorilla. | 10–8 |
| Subtribe | Hominina | Genus Homo and close human relatives and ancestors after splitting from Pan—the hominins | 8–4 |
| (Genus) | Ardipithecus s.l. | 6-4 | |
| (Genus) | Australopithecus | 3 | |
| Genus | Homo (H. habilis) | Humans | 2.5 |
| (Species) | H. erectus s.l. | ||
| (Species) | H. heidelbergensis s.l. | ||
| Species | Homo sapiens s.s. | Anatomically modern humans | 0.8–0.3 |
| Date | Event |
|---|---|
| 4.3-4.1 Ga | The earliest life appears, possibly as protocells. Their genetic material was probably composed of RNA, capable of both self replication and enzymatic activity; their membranes were composed of lipids. The genes were separate strands, translated into proteins and often exchanged between the protocells. Further information: Abiogenesis, RNA world, and Earliest known life forms |
| 4.0-3.8 Ga | Prokaryotic cells appear; their genetic materials are composed of the more stable DNA and they use proteins for various reasons, primarily for aiding DNA to replicate itself by proteinaceous enzymes (RNA now acts as an intermediary in this central dogma of genetic information flow of cellular life); genes are now linked in sequences so all information passes to offsprings. They had cell walls & outer membranes and were probably initially thermophiles. Further information: Cell (biology) § Origins |
| 3.5 Ga | This marks the first appearance of cyanobacteria and their method of oxygenic photosynthesis and therefore the first occurrence of atmospheric oxygen on Earth. For another billion years, prokaryotes would continue to diversify undisturbed. Further information: Evolution of photosynthesis § Origin, and Great Oxidation Event |
| 2.5-2.2 Ga | First organisms to use oxygen. By 2400 Ma, in what is referred to as the Great Oxidation Event, (GOE), most of the pre-oxygen anaerobic forms of life were wiped out by the oxygen producers. Further information: Geological history of oxygen |
| 2.2-1.8 Ga | Origin of the eukaryotes: organisms with nuclei, endomembrane systems (including mitochondria) and complex cytoskeletons; they spliced mRNA between transcription and translation (splicing also occurs in prokaryotes, but it is only of non-coding RNAs). The evolution of eukaryotes, and possibly sex, is thought to be related to the GOE, as it probably pressured two or three lineages of prokaryotes (including an aerobe one, which later became mitochondria) to depend on each other, leading to endosymbiosis. Early eukaryotes lost their cell walls and outer membranes. Further information: Eukaryote § Origin of eukaryotes |
| 1.2 Ga | Sexual reproduction evolves (mitosis and meiosis) by this time at least, leading to faster evolution where genes are mixed in every generation enabling greater variation for subsequent selection. |
| 1.2-0.8 Ga |  The Holozoa lineage of eukaryotes evolves many features for making cell colonies, and finally leads to the ancestor of animals (metazoans) and choanoflagellates. Proterospongia (members of the Choanoflagellata) are the best living examples of what the ancestor of all animals may have looked like. They live in colonies, and show a primitive level of cellular specialization for different tasks. |
Animalia
| Date | Event |
|---|---|
| 800–650 Ma |  Urmetazoan: The first fossils that might represent animals appear in the 665-million-year-old rocks of the Trezona Formation of South Australia. These fossils are interpreted as being early sponges. Multicellular animals may have existed from 800 Ma. Separation from the Porifera (sponges) lineage. Eumetazoa/Diploblast: separation from the Ctenophora ("comb jellies") lineage. Planulozoa/ParaHoxozoa: separation from the Placozoa and Cnidaria lineages. All diploblasts possess epithelia, nerves, muscles and connective tissue and mouths, and except for placozoans, have some form of symmetry, with their ancestors probably having radial symmetry like that of cnidarians. Diploblasts separated their early embryonic cells into two germ layers (ecto- and endoderm). Photoreceptive eye-spots evolve. |
| 650-600 Ma |  Urbilaterian: the last common ancestor of xenacoelomorphs, protostomes (including the arthropod [insect, crustacean, spider], mollusc [squid, snail, clam] and annelid [earthworm] lineages) and the deuterostomes (including the vertebrate [human] lineage) (the last two are more related to each other and called Nephrozoa). Xenacoelomorphs all have a gonopore to expel gametes but nephrozoans merged it with their anus. Earliest development of bilateral symmetry, mesoderm, head (anterior cephalization) and various gut muscles (and thus peristalsis) and, in the Nephrozoa, nephridia (kidney precursors), coelom (or maybe pseudocoelom), distinct mouth and anus (evolution of through-gut), and possibly even nerve cords and blood vessels. Reproductive tissue probably concentrates into a pair of gonads connecting just before the posterior orifice. "Cup-eyes" and balance organs evolve (the function of hearing added later as the more complex inner ear evolves in vertebrates). The nephrozoan through-gut had a wider portion in the front, called the pharynx. The integument or skin consists of an epithelial layer (epidermis) and a connective layer. |
| 600-540 Ma |  Most known animal phyla appeared in the fossil record as marine species during the Ediacaran-Cambrian explosion, probably caused by long scale oxygenation since around 585 Ma (sometimes called the Neoproterozoic Oxygenation Event or NOE) and also an influx of oceanic minerals. Deuterostomes, the last common ancestor of the Chordata [human] lineage, Hemichordata (acorn worms and graptolites) and Echinodermata (starfish, sea urchins, sea cucumbers, etc.), probably had both ventral and dorsal nerve cords like modern acorn worms. An archaic survivor from this stage is the acorn worm, sporting an open circulatory system (with less branched blood vessels) with a heart that also functions as a kidney. Acorn worms have a plexus concentrated into both dorsal and ventral nerve cords. The dorsal cord reaches into the proboscis, and is partially separated from the epidermis in that region. This part of the dorsal nerve cord is often hollow, and may well be homologous with the brain of vertebrates. Deuterostomes also evolved pharyngeal slits, which were probably used for filter feeding like in hemi- and proto-chordates. |
Chordata
| Date | Event |
|---|---|
| 540-520 Ma |  The increased amount of oxygen causes many eukaryotes, including most animals, to become obligate aerobes. The Chordata ancestor gave rise to the lancelets (Amphioxii) and Olfactores. Ancestral chordates evolved a post-anal tail, notochord, and endostyle (precursor of thyroid). The pharyngeal slits (or gills) are now supported by connective tissue and used for filter feeding and possibly breathing. The first of these basal chordates to be discovered by science was Pikaia gracilens. Other, earlier chordate predecessors include Myllokunmingia fengjiaoa, Yunnanozoon lividum, and Haikouichthys ercaicunensis. They probably lost their ventral nerve cord and evolved a special region of the dorsal one, called the brain, with glia becoming permanently associated with neurons. They probably evolved the first blood cells (probably early leukocytes, indicating advanced innate immunity), which they made around the pharynx and gut. All chordates except tunicates sport an intricate, closed circulatory system, with highly branched blood vessels. Olfactores, last common ancestor of tunicates and vertebrates in which olfaction (smell) evolved. Since lancelets lack a heart, it possibly emerged in this ancestor (previously the blood vessels themselves were contractile) though it could have been lost in lancelets after evolving in early deuterostomes (hemichordates and echinoderms have hearts). |
| 520-480 Ma |  The first vertebrates ("fish") appear: the Agnathans. They were jawless, had seven pairs of pharyngeal arches like their descendants today, and their endoskeletons were cartilaginous (then only consisting of the chondrocranium/braincase and vertebrae). The jawless Cyclostomata diverge at this stage. The connective tissue below the epidermis differentiates into the dermis and hypodermis. They depended on gills for respiration and evolved the unique sense of taste (the remaining sense of the skin now called "touch"), endothelia, camera eyes and inner ears (capable of hearing and balancing; each consists of a lagena, an otolithic organ and two semicircular canals) as well as livers, thyroids, kidneys and two-chambered hearts (one atrium and one ventricle). They had a tail fin but lacked the paired (pectoral and pelvic) fins of more advanced fish. Brain divided into three parts (further division created distinct regions based on function). The pineal gland of the brain penetrates to the level of the skin on the head, making it seem like a third eye. They evolved the first erythrocytes and thrombocytes. |
| 460-430 Ma |  The Placodermi were the first jawed fishes (Gnathostomata); their jaws evolved from the first gill/pharyngeal arch and they largely replaced their endoskeletal cartilage with bone and evolved pectoral and pelvic fins. Bones of the first gill arch became the upper and lower jaw, while those from the second arch became the hyomandibula, ceratohyal and basihyal; this closed two of the seven pairs of gills. The gap between the first and second arches just below the braincase (fused with upper jaw) created a pair of spiracles, which opened in the skin and led to the pharynx (water passed through them and left through gills). Placoderms had competition with the previous dominant animals, the cephalopods and sea scorpions, and rose to dominance themselves. A lineage of them probably evolved into the bony and cartilaginous fish, after evolving scales, teeth (which allowed the transition to full carnivory), stomachs, spleens, thymuses, myelin sheaths, hemoglobin and advanced, adaptive immunity (the latter two occurred independently in the lampreys and hagfish). Jawed fish also have a third, lateral semicircular canal and their otoliths are divided between a saccule and utricle. |
| 430-410 Ma |  |
Tetrapoda
Further information: Evolution of tetrapods
| Date | Event |
|---|---|
| 390 Ma |  Some freshwater lobe-finned fish (sarcopterygii) develop limbs and give rise to the Tetrapodomorpha. These fish evolved in shallow and swampy freshwater habitats, where they evolved large eyes and spiracles. Primitive tetrapods ("fishapods") developed from tetrapodomorphs with a two-lobed brain in a flattened skull, a wide mouth and a medium snout, whose upward-facing eyes show that it was a bottom-dweller, and which had already developed adaptations of fins with fleshy bases and bones. (The "living fossil" coelacanth is a related lobe-finned fish without these shallow-water adaptations.) Tetrapod fishes used their fins as paddles in shallow-water habitats choked with plants and detritus. The universal tetrapod characteristics of front limbs that bend backward at the elbow and hind limbs that bend forward at the knee can plausibly be traced to early tetrapods living in shallow water. Panderichthys is a 90–130 cm (35–50 in) long fish from the Late Devonian period (380 Mya). It has a large tetrapod-like head. Panderichthys exhibits features transitional between lobe-finned fishes and early tetrapods. Trackway impressions made by something that resembles Ichthyostega's limbs were formed 390 Ma in Polish marine tidal sediments. This suggests tetrapod evolution is older than the dated fossils of Panderichthys through to Ichthyostega. |
| 375-350 Ma |  Tiktaalik is a genus of sarcopterygian (lobe-finned) fishes from the late Devonian with many tetrapod-like features. It shows a clear link between Panderichthys and Acanthostega.   Acanthostega is an extinct tetrapod, among the first animals to have recognizable limbs. It is a candidate for being one of the first vertebrates to be capable of coming onto land. It lacked wrists, and was generally poorly adapted for life on land. The limbs could not support the animal's weight. Acanthostega had both lungs and gills, also indicating it was a link between lobe-finned fish and terrestrial vertebrates. The dorsal pair of ribs form a rib cage to support the lungs, while the ventral pair disappears. Ichthyostega is another extinct tetrapod. Being one of the first animals with only two pairs of limbs (also unique since they end in digits and have bones), Ichthyostega is seen as an intermediate between a fish and an amphibian. Ichthyostega had limbs but these probably were not used for walking. They may have spent very brief periods out of water and would have used their limbs to paw their way through the mud. They both had more than five digits (eight or seven) at the end of each of their limbs, and their bodies were scaleless (except their bellies, where they remained as gastralia). Many evolutionary changes occurred at this stage: eyelids and tear glands evolved to keep the eyes wet out of water and the eyes became connected to the pharynx for draining the liquid; the hyomandibula (now called columella) shrank into the spiracle, which now also connected to the inner ear at one side and the pharynx at another, becoming the Eustachian tube (columella assisted in hearing); an early eardrum (a patch of connective tissue) evolved on the end of each tube (called the otic notch); and the ceratohyal and basihyal merged into the hyoid. These "fishapods" had more ossified and stronger bones to support themselves on land (especially skull and limb bones). Jaw bones fuse together while gill and opercular bones disappear. |
| 350-330 Ma |  Pederpes from around 350 Ma indicates that the standard number of 5 digits evolved at the Early Carboniferous, when modern tetrapods (or "amphibians") split in two directions (one leading to the extant amphibians and the other to amniotes). At this stage, our ancestors evolved vomeronasal organs, salivary glands, tongues, parathyroid glands, three-chambered hearts (with two atria and one ventricle) and bladders, and completely removed their gills by adulthood. The glottis evolves to prevent food going into the respiratory tract. Lungs and thin, moist skin allowed them to breathe; water was also needed to give birth to shell-less eggs and for early development. Dorsal, anal and tail fins all disappeared. Lissamphibia (extant amphibians) retain many features of early amphibians but they have only four digits (caecilians have none). |
| 330-300 Ma |  From amphibians came the first amniotes: Hylonomus, a primitive reptile, is the earliest amniote known. It was 20 cm (8 in) long (including the tail) and probably would have looked rather similar to modern lizards. It had small sharp teeth and probably ate small millipedes and insects. It is a precursor of later amniotes (including both the reptiles and the ancestors of mammals). Alpha keratin first evolves here; it is used in the claws of modern amniotes, and hair in mammals, indicating claws and a different type of scales evolved in amniotes (complete loss of gills as well). Evolution of the amniotic egg allows the amniotes to reproduce on land and lay shelled eggs on dry land. They did not need to return to water for reproduction nor breathing. This adaptation and the desiccation-resistant scales gave them the capability to inhabit the uplands for the first time, albeit making them drink water through their mouths. At this stage, adrenal tissue may have concentrated into discrete glands. Amniotes have advanced nervous systems, with twelve pairs of cranial nerves, unlike lower vertebrates. They also evolved true sternums but lost their eardrums and otic notches (hearing only by columella bone conduction). |
Mammalia
Further information: Evolution of mammals
| Date | Event |
|---|---|
| 300-260 Ma | Shortly after the appearance of the first amniotes, two branches split off. One branch is the Sauropsida, from which come the reptiles, including birds. The other branch is Synapsida, from which come modern mammals. Both had temporal fenestrae, a pair of holes in their skulls behind the eyes, which were used to increase the space for jaw muscles. Synapsids had one opening on each side, while diapsids (a branch of Sauropsida) had two. An early, inefficient version of diaphragm may have evolved in synapsids. The earliest synapsids, or "proto-mammals," are the pelycosaurs. The pelycosaurs were the first animals to have temporal fenestrae. Pelycosaurs were not therapsids but their ancestors. The therapsids were, in turn, the ancestors of mammals. The therapsids had temporal fenestrae larger and more mammal-like than pelycosaurs, their teeth showed more serial differentiation, their gait was semi-erect and later forms had evolved a secondary palate. A secondary palate enables the animal to eat and breathe at the same time and is a sign of a more active, perhaps warm-blooded, way of life. They had lost gastralia and, possibly, scales. |
| 260-230 Ma |  One subgroup of therapsids, the cynodonts, lose pineal eye and lumbar ribs and very likely became warm-blooded. The lower respiratory tract forms intricate branches in the lung parenchyma, ending in highly vascularized alveoli. Erythrocytes and thrombocytes lose their nuclei while lymphatic systems and advanced immunity emerge. They may have also had thicker dermis like mammals today. The jaws of cynodonts resembled modern mammal jaws; the anterior portion, the dentary, held differentiated teeth. This group of animals likely contains a species which is the ancestor of all modern mammals. Their temporal fenestrae merged with their orbits. Their hindlimbs became erect and their posterior bones of the jaw progressively shrunk to the region of the columella. |
| 230-170 Ma | 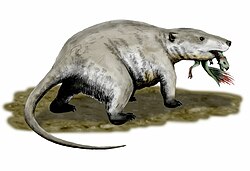 From Eucynodontia came the first mammals. Most early mammals were small shrew-like animals that fed on insects and had transitioned to nocturnality to avoid competition with the dominant archosaurs — this led to the loss of the vision of red and ultraviolet light (ancestral tetrachromacy of vertebrates reduced to dichromacy). Although there is no evidence in the fossil record, it is likely that these animals had a constant body temperature, hair and milk glands for their young (the glands stemmed from the milk line). The neocortex (part of the cerebrum) region of the brain evolves in Mammalia, at the reduction of the tectum (non-smell senses which were processed here became integrated into neocortex but smell became primary sense). Origin of the prostate gland and a pair of holes opening to the columella and nearby shrinking jaw bones; new eardrums stand in front of the columella and Eustachian tube. The skin becomes hairy, glandular (glands secreting sebum and sweat) and thermoregulatory. Teeth fully differentiate into incisors, canines, premolars and molars; mammals become diphyodont and possess developed diaphragms and males have internal penises. All mammals have four chambered hearts (with two atria and two ventricles) and lack cervical ribs (now mammals only have thoracic ribs). Monotremes are an egg-laying group of mammals represented today by the platypus and echidna. Recent genome sequencing of the platypus indicates that its sex genes are closer to those of birds than to those of the therian (live birthing) mammals. Comparing this to other mammals, it can be inferred that the first mammals to gain sexual differentiation through the existence or lack of SRY gene (found in the y-Chromosome) evolved only in the therians. Early mammals and possibly their eucynodontian ancestors had epipubic bones, which serve to hold the pouch in modern marsupials (in both sexes). |
| 170-120 Ma |  Evolution of live birth (viviparity), with early therians probably having pouches for keeping their undeveloped young like in modern marsupials. Nipples stemmed out of the therian milk lines. The posterior orifice separates into anal and urogenital openings; males possess an external penis. Monotremes and therians independently detach the malleus and incus from the dentary (lower jaw) and combine them to the shrunken columella (now called stapes) in the tympanic cavity behind the eardrum (which is connected to the malleus and held by another bone detached from the dentary, the tympanic plus ectotympanic), and coil their lagena (cochlea) to advance their hearing, with therians further evolving an external pinna and erect forelimbs. Female placentalian mammals do not have pouches and epipubic bones but instead have a developed placenta which penetrates the uterus walls (unlike marsupials), allowing a longer gestation; they also have separated urinary and genital openings. |
| 100-90 Ma | Last common ancestor of rodents, rabbits, ungulates, carnivorans, bats, shrews and humans (base of the clade Boreoeutheria; males now have external testicles). |
Primates
Further information: Evolution of primates
| Date | Event |
|---|---|
| 90–66 Ma |   A group of small, nocturnal, arboreal, insect-eating mammals called Euarchonta begins a speciation that will lead to the orders of primates, treeshrews and flying lemurs. They reduced the number of mammaries to only two pairs (on the chest). Primatomorpha is a subdivision of Euarchonta including primates and their ancestral stem-primates Plesiadapiformes. An early stem-primate, Plesiadapis, still had claws and eyes on the side of the head, making it faster on the ground than in the trees, but it began to spend long times on lower branches, feeding on fruits and leaves. The Plesiadapiformes very likely contain the ancestor species of all primates. They first appeared in the fossil record around 66 million years ago, soon after the Cretaceous–Paleogene extinction event that eliminated about three-quarters of plant and animal species on Earth, including most dinosaurs. One of the last Plesiadapiformes is Carpolestes simpsoni, having grasping digits but not forward-facing eyes. |
| 66-56 Ma | Primates diverge into suborders Strepsirrhini (wet-nosed primates) and Haplorrhini (dry-nosed primates). Brain expands and cerebrum divides into 4 pairs of lobes. The postorbital bar evolves to separate the orbit from the temporal fossae as sight regains its position as the primary sense; eyes became forward-facing. Strepsirrhini contain most prosimians; modern examples include lemurs and lorises. The haplorrhines include the two living groups: prosimian tarsiers, and simian monkeys, including apes. The Haplorrhini metabolism lost the ability to produce vitamin C, forcing all descendants to include vitamin C-containing fruit in their diet. Early primates only had claws in their second digits; the rest were turned into nails. |
| 50-35 Ma |  Simians split into infraorders Platyrrhini and Catarrhini. They fully transitioned to diurnality and lacked any claw and tapetum lucidum (which evolved many times in various vertebrates). They possibly evolved at least some of the paranasal sinuses, and transitioned from estrous cycle to menstrual cycle. The number of mammaries is now reduced to only one thoracic pair. Platyrrhines, New World monkeys, have prehensile tails and males are color blind. The individuals whose descendants would become Platyrrhini are conjectured to have migrated to South America either on a raft of vegetation or via a land bridge (the hypothesis now favored). Catarrhines mostly stayed in Africa as the two continents drifted apart. Possible early ancestors of catarrhines include Aegyptopithecus and Saadanius. |
| 35-20 Ma |  Catarrhini splits into 2 superfamilies, Old World monkeys (Cercopithecoidea) and apes (Hominoidea). Human trichromatic color vision had its genetic origins in this period. Catarrhines lost the vomeronasal organ (or possibly reduced it to vestigial status). Proconsul was an early genus of catarrhine primates. They had a mixture of Old World monkey and ape characteristics. Proconsul's monkey-like features include thin tooth enamel, a light build with a narrow chest and short forelimbs, and an arboreal quadrupedal lifestyle. Its ape-like features are its lack of a tail, ape-like elbows, and a slightly larger brain relative to body size. Proconsul africanus is a possible ancestor of both great and lesser apes, including humans. |
Hominidae
| Date | Event |
|---|---|
| 20-15 Ma | Hominidae (great ape ancestors) speciate from the ancestors of the gibbon (lesser apes) between c. 20 to 16 Ma. They largely reduced their ancestral snout and lost the uricase enzyme (present in most organisms). |
| 16-12 Ma | Homininae ancestors speciate from the ancestors of the orangutan between c. 18 to 14 Ma. Pierolapithecus catalaunicus is thought to be a common ancestor of humans and the other great apes, or at least a species that brings us closer to a common ancestor than any previous fossil discovery. It had the special adaptations for tree climbing as do present-day humans and other great apes: a wide, flat rib cage, a stiff lower spine, flexible wrists, and shoulder blades that lie along its back. |
| 12 Ma | Danuvius guggenmosi is the first-discovered Late Miocene great ape with preserved long bones, and greatly elucidates the anatomical structure and locomotion of contemporary apes. It had adaptations for both hanging in trees (suspensory behavior) and walking on two legs (bipedalism)—whereas, among present-day hominids, humans are better adapted for the latter and the others for the former. Danuvius thus had a method of locomotion unlike any previously known ape called "extended limb clambering", walking directly along tree branches as well as using arms for suspending itself. The last common ancestor between humans and other apes possibly had a similar method of locomotion. |
| 12-8 Ma | The clade currently represented by humans and the genus Pan (chimpanzees and bonobos) splits from the ancestors of the gorillas between c. 12 to 8 Ma. |
| 8-6 Ma |  Hominini: The latest common ancestor of humans and chimpanzees is estimated to have lived between roughly 10 to 5 million years ago. Both chimpanzees and humans have a larynx that repositions during the first two years of life to a spot between the pharynx and the lungs, indicating that the common ancestors have this feature, a precondition for vocalized speech in humans. Speciation may have begun shortly after 10 Ma, but late admixture between the lineages may have taken place until after 5 Ma. Candidates of Hominina or Homininae species which lived in this time period include Graecopithecus (c. 7 Ma), Sahelanthropus tchadensis (c. 7 Ma), Orrorin tugenensis (c. 6 Ma).  Ardipithecus was arboreal, meaning it lived largely in the forest where it competed with other forest animals for food, no doubt including the contemporary ancestor of the chimpanzees. Ardipithecus was probably bipedal as evidenced by its bowl shaped pelvis, the angle of its foramen magnum and its thinner wrist bones, though its feet were still adapted for grasping rather than walking for long distances. |
| 4-3.5 Ma |  A member of the Australopithecus afarensis left human-like footprints on volcanic ash in Laetoli, northern Tanzania, providing strong evidence of full-time bipedalism. Australopithecus afarensis lived between 3.9 and 2.9 million years ago, and is considered one of the earliest hominins—those species that developed and comprised the lineage of Homo and Homo's closest relatives after the split from the line of the chimpanzees. It is thought that A. afarensis was ancestral to both the genus Australopithecus and the genus Homo. Compared to the modern and extinct great apes, A. afarensis had reduced canines and molars, although they were still relatively larger than in modern humans. A. afarensis also has a relatively small brain size (380–430 cm3) and a prognathic (anterior-projecting) face. Australopithecines have been found in savannah environments; they probably developed their diet to include scavenged meat. Analyses of Australopithecus africanus lower vertebrae suggests that these bones changed in females to support bipedalism even during pregnancy. |
| 3.5–3.0 Ma | Kenyanthropus platyops, a possible ancestor of Homo, emerges from the Australopithecus. Stone tools are deliberately constructed, possibly by Kenyanthropus platyops or Australopithecus afarensis. |
| 3 Ma | The bipedal australopithecines (a genus of the subtribe Hominina) evolve in the savannas of Africa being hunted by Megantereon. Loss of body hair occurs from 3 to 2 Ma, in parallel with the development of full bipedalism and slight enlargement of the brain. |
Homo
| Date | Event |
|---|---|
| 2.8–2.0 Ma | Early Homo appears in East Africa, speciating from australopithecine ancestors. The Lower Paleolithic is defined by the beginning of use of stone tools. Australopithecus garhi was using stone tools at about 2.5 Ma. Homo habilis is the oldest species given the designation Homo, by Leakey et al. in 1964. H. habilis is intermediate between Australopithecus afarensis and H. erectus, and there have been suggestions to re-classify it within genus Australopithecus, as Australopithecus habilis. LD 350-1 is now considered the earliest known specimen of the genus Homo, dating to 2.75–2.8 Ma, found in the Ledi-Geraru site in the Afar Region of Ethiopia. It is currently unassigned to a species, and it is unclear if it represents the ancestor to H. habilis and H. rudolfensis, which are estimated to have evolved around 2.4 Ma. Stone tools found at the Shangchen site in China and dated to 2.12 million years ago are considered the earliest known evidence of hominins outside Africa, surpassing Dmanisi hominins found in Georgia by 300,000 years, although whether these hominins were an early species in the genus Homo or another hominin species is unknown. Further information: Homo naledi and Homo rudolfensis |
| 1.9–0.8 Ma |  Homo erectus derives from early Homo or late Australopithecus. Homo habilis, although significantly different of anatomy and physiology, is thought to be the ancestor of Homo ergaster, or African Homo erectus; but it is also known to have coexisted with H. erectus for almost half a million years (until about 1.5 Ma). From its earliest appearance at about 1.9 Ma, H. erectus is distributed in East Africa and Southwest Asia (Homo georgicus). H. erectus is the first known species to develop control of fire, by about 1.5 Ma. H. erectus later migrates throughout Eurasia, reaching Southeast Asia by 0.7 Ma. It is described in a number of subspecies. Early humans were social and initially scavenged, before becoming active hunters. The need to communicate and hunt prey efficiently in a new, fluctuating environment (where the locations of resources need to be memorized and told) may have driven the expansion of the brain from 2 to 0.8 Ma. Evolution of dark skin at about 1.2 Ma. Homo antecessor may be a common ancestor of Homo sapiens and Neanderthals. At present estimate, humans have approximately 20,000–25,000 genes and share 99% of their DNA with the now extinct Neanderthal and 95–99% of their DNA with their closest living evolutionary relative, the chimpanzees. The human variant of the FOXP2 gene (linked to the control of speech) has been found to be identical in Neanderthals. |
| 0.8–0.3 Ma |  Divergence of Neanderthal and Denisovan lineages from a common ancestor. Homo heidelbergensis (in Africa also known as Homo rhodesiensis) had long been thought to be a likely candidate for the last common ancestor of the Neanderthal and modern human lineages. However, genetic evidence from the Sima de los Huesos fossils published in 2016 seems to suggest that H. heidelbergensis in its entirety should be included in the Neanderthal lineage, as "pre-Neanderthal" or "early Neanderthal", while the divergence time between the Neanderthal and modern lineages has been pushed back to before the emergence of H. heidelbergensis, to about 600,000 to 800,000 years ago, the approximate age of Homo antecessor. Brain expansion (enlargement) between 0.8 and 0.2 Ma may have occurred due to the extinction of most African megafauna (which made humans feed from smaller prey and plants, which required greater intelligence due to greater speed of the former and uncertainty about whether the latter were poisonous or not), extreme climate variability after Mid-Pleistocene Transition (which intensified the situation, and resulted in frequent migrations), and in general selection for more social life (and intelligence) for greater chance of survival, reproductivity, and care for mothers. Solidified footprints dated to about 350 ka and associated with H. heidelbergensis were found in southern Italy in 2003. H. sapiens lost the brow ridges from their hominid ancestors as well as the snout completely, though their noses evolve to be protruding (possibly from the time of H. erectus). By 200 ka, humans had stopped their brain expansion. |
Homo sapiens
Further information: Homo sapiens, Neanderthal, Interbreeding between archaic and modern humans, Recent human evolution, and Human genetic variation
| Date | Event |
|---|---|
| 300–130 ka | .jpg/250px-Homo_sapiens_sapiens_(Fundort_Jebel_Irhoud_Marokko).jpg) Neanderthals and Denisovans emerge from the northern Homo heidelbergensis lineage around 500-450 ka while sapients emerge from the southern lineage around 350-300 ka. Fossils attributed to H. sapiens, along with stone tools, dated to approximately 300,000 years ago, found at Jebel Irhoud, Morocco yield the earliest fossil evidence for anatomically modern Homo sapiens. Modern human presence in East Africa (Gademotta), at 276 kya. In July 2019, anthropologists reported the discovery of 210,000 year old remains of what may possibly have been a H. sapiens in Apidima Cave, Peloponnese, Greece. Patrilineal and matrilineal most recent common ancestors (MRCAs) of living humans roughly between 200 and 100 kya with some estimates on the patrilineal MRCA somewhat higher, ranging up to 250 to 500 kya. 160,000 years ago, Homo sapiens idaltu in the Awash River Valley (near present-day Herto village, Ethiopia) practiced excarnation. |
| 130–80 ka | Marine Isotope Stage 5 (Eemian). Modern human presence in Southern Africa and West Africa. Appearance of mitochondrial haplogroup (mt-haplogroup) L2. |
| 80–50 ka | MIS 4, beginning of the Upper Paleolithic. Early evidence for behavioral modernity. Appearance of mt-haplogroups M and N. Southern Dispersal migration out of Africa, Proto-Australoid peopling of Oceania. Archaic admixture from Neanderthals in Eurasia, from Denisovans in Oceania with trace amounts in Eastern Eurasia, and from an unspecified African lineage of archaic humans in Sub-Saharan Africa as well as an interbred species of Neanderthals and Denisovans in Asia and Oceania. |
| 50–25 ka |
.jpg/250px-Homo_sapiens_sapiens,_Oase,_Rumänien_(Daniela_Hitzemann).jpg) Behavioral modernity develops by this time or earlier, according to the "great leap forward" theory. Extinction of Homo floresiensis. M168 mutation (carried by all non-African males). Appearance of mt-haplogroups U and K. Peopling of Europe, peopling of the North Asian Mammoth steppe. Paleolithic art. Extinction of Neanderthals and other archaic human variants (with possible survival of hybrid populations in Asia and Africa). Appearance of Y-Haplogroup R2; mt-haplogroups J and X. |
| after 25 ka |  Last Glacial Maximum; Epipaleolithic / Mesolithic / Holocene. Peopling of the Americas. Appearance of: Y-Haplogroup R1a; mt-haplogroups V and T. Various recent divergence associated with environmental pressures, e.g. light skin in Europeans and East Asians (KITLG, ASIP), after 30 ka; Inuit adaptation to high-fat diet and cold climate, 20 ka. Extinction of late surviving archaic humans at the beginning of the Holocene (12 ka). Accelerated divergence due to selection pressures in populations participating in the Neolithic Revolution after 12 ka, e.g. East Asian types of ADH1B associated with rice domestication, or lactase persistence. A slight decrease in brain size occurred a few thousand years ago.[citation needed] |
The earliest Solar System
|
Nature timeline
|
||||||||||||||||||||||||||||||||||||||
|
−13 —
–
−12 —
–
−11 —
–
−10 —
–
−9 —
–
−8 —
–
−7 —
–
−6 —
–
−5 —
–
−4 —
–
−3 —
–
−2 —
–
−1 —
–
0 —
|
Dark Ages
Reionization
Matter-dominated
era Water on Earth
Life
Multicellular
life Vertebrates
|
|
||||||||||||||||||||||||||||||||||||
|
(billion years ago)
|
||||||||||||||||||||||||||||||||||||||
Main articles: Formation and evolution of the Solar System and Nebular hypothesis
In the earliest Solar System history, the Sun, the planetesimals and the giant planets were formed. The inner Solar System aggregated more slowly than the outer, so the terrestrial planets were not yet formed, including Earth and Moon.
- c. 4,570 Ma – A supernova explosion (known as the primal supernova) seeds our galactic neighborhood with heavy elements that will be incorporated into the Earth, and results in a shock wave in a dense region of the Milky Way galaxy. The Ca-Al-rich inclusions, which formed 2 million years before the chondrules, are a key signature of a supernova explosion.
- c. 4,567 ±3 Ma – Rapid collapse of hydrogen molecular cloud, forming a third-generation Population I star, the Sun, in a region of the Galactic Habitable Zone (GHZ), about 25,000 light years from the center of the Milky Way Galaxy.
- c. 4,566 ±2 Ma – A protoplanetary disc (from which Earth eventually forms) emerges around the young Sun, which is in its T Tauri stage.
- c. 4,560–4,550 Ma – Proto-Earth forms at the outer (cooler) edge of the habitable zone of the Solar System. At this stage the solar constant of the Sun was only about 73% of its current value, but liquid water may have existed on the surface of the Proto-Earth, probably due to the greenhouse warming of high levels of methane and carbon dioxide present in the atmosphere. Early bombardment phase begins: because the solar neighbourhood is rife with large planetoids and debris, Earth experiences a number of giant impacts that help to increase its overall size.
Precambrian Supereon
Main article: Precambrian
- c. 4,533 Ma – The Precambrian (to c. 539 Ma), now termed a "supereon" but formerly an era, is split into three geological time intervals called eons: Hadean, Archaean and Proterozoic. The latter two are sub-divided into several eras as currently defined. In total, the Precambrian comprises some 85% of geological time from the formation of Earth to the time when creatures first developed exoskeletons (i.e., hard outer parts) and thereby left abundant fossil remains.
Hadean Eon
Main article: Hadean
- c. 4,533 Ma – Hadean Eon, Precambrian Supereon and unofficial Cryptic era start as the Earth–Moon system forms, possibly as a result of a glancing collision between proto-Earth and the hypothetical protoplanet Theia (the Earth was considerably smaller than now, before this impact). This impact vaporized a large amount of the crust, and sent material into orbit around Earth, which lingered as rings, similar to those of Saturn, for a few million years, until they coalesced to become the Moon. The Moon geology pre-Nectarian period starts. Earth was covered by a magmatic ocean 200 kilometres (120 mi) deep resulting from the impact energy from this and other planetesimals during the early bombardment phase, and energy released by the planetary core forming. Outgassing from crustal rocks gives Earth a reducing atmosphere of methane, nitrogen, hydrogen, ammonia, and water vapour, with lesser amounts of hydrogen sulfide, carbon monoxide, then carbon dioxide. With further full outgassing over 1000–1500 K, nitrogen and ammonia become lesser constituents, and comparable amounts of methane, carbon monoxide, carbon dioxide, water vapour, and hydrogen are released.
- c. 4,500 Ma – Sun enters main sequence: a solar wind sweeps the Earth-Moon system clear of debris (mainly dust and gas). End of the Early Bombardment Phase. Basin Groups Era begins on Earth.
- c. 4,450 Ma – 100 million years after the Moon formed, the first lunar crust, formed of lunar anorthosite, differentiates from lower magmas. The earliest Earth crust probably forms similarly out of similar material. On Earth the pluvial period starts, in which the Earth's crust cools enough to let oceans form.
- c. 4,404 Ma – First known mineral, found at Jack Hills in Western Australia. Detrital zircons show presence of a solid crust and liquid water. Latest possible date for a secondary atmosphere to form, produced by the Earth's crust outgassing, reinforced by water and possibly organic molecules delivered by comet impacts and carbonaceous chondrites (including type CI shown to be high in a number of amino acids and polycyclic aromatic hydrocarbons (PAH)).
- c. 4,300 Ma – Nectarian Era begins on Earth.
- c. 4,250 Ma – Earliest evidence for life, based on unusually high amounts of light isotopes of carbon, a common sign of life, found in Earth's oldest mineral deposits located in the Jack Hills of Western Australia.
- c. 4,100 Ma – Early Imbrian Era begins on Earth. Late heavy bombardment of the Moon (and probably of the Earth as well) by bolides and asteroids, produced possibly by the planetary migration of Neptune into the Kuiper belt as a result of orbital resonances between Jupiter and Saturn. "Remains of biotic life" were found in 4.1 billion-year-old rocks in Western Australia. According to one of the researchers, "If life arose relatively quickly on Earth ... then it could be common in the universe."
Archean Eon
Main article: Archean
Eoarchean Era
Main article: Eoarchean
- c. 4,031 Ma – Archean Eon and Eoarchean Era start. Possible first appearance of plate tectonic activity in the Earth's crust as plate structures may have begun appearing. Possible beginning of Napier Mountains Orogeny forces of faulting and folding create first metamorphic rocks. Origins of life.
- c. 4,030 Ma – Acasta Gneiss of Northwest Territories, Canada, first known oldest rock, or aggregate of minerals.
- c. 3,930 Ma – Possible stabilization of Canadian Shield begins
- c. 3,920–3,850 Ma – Final phase of Late Heavy Bombardment
- c. 3,850 Ma – Greenland apatite shows evidence of 12C enrichment, characteristic of the presence of photosynthetic life.
- c. 3,850 Ma – Evidence of life: Akilia Island graphite off Western Greenland contains evidence of kerogen, of a type consistent with photosynthesis.[citation needed]
- c. 3,800 Ma – Oldest banded iron formations found. First complete continental masses or cratons, formed of granite blocks, appear on Earth. Occurrence of initial felsic igneous activity on eastern edge of Antarctic craton as first great continental mass begins to coalesce. East European Craton begins to form – first rocks of the Ukrainian Shield and Voronezh Massif are laid down
- c. 3,750 Ma – Nuvvuagittuq Greenstone Belt forms
- c. 3,700 Ma – Graphite found to be biogenic in 3.7 billion-year-old metasedimentary rocks discovered in Western Greenland Stabilization of Kaapval craton begins: old tonaltic gneisses laid down
Paleoarchean Era
- c. 3,600 Ma – Paleoarchean Era starts. Possible assembly of the Vaalbara supercontinent; oldest cratons on Earth (such as the Canadian Shield, East European Craton and Kaapval) begin growing as a result of crustal disturbances along continents coalescing into Vaalbara – Pilbara Craton stabilizes. Formation of Barberton greenstone belt: Makhonjwa Mountains uplifts on the eastern edge of Kaapval craton, oldest mountains in Africa – area called the "genesis of life" for exceptional preservation of fossils. Narryer gneiss terrane stabilizes: these gneisses become the "bedrock" for the formation of the Yilgarn craton in Australia – noted for the survival of the Jack Hills where the oldest mineral, a zircon was uncovered.
- c. 3,500 Ma – Lifetime of the Last universal ancestor: split between bacteria and archaea occurs as "tree of life" begins branching out – varieties of Eubacteria begin to radiate out globally. Fossils resembling cyanobacteria, found at Warrawoona, Western Australia.[citation needed]
- c. 3,480 Ma – Fossils of microbial mat found in 3.48 billion-year-old sandstone discovered in Western Australia. First appearance of stromatolitic organisms that grow at interfaces between different types of material, mostly on submerged or moist surfaces.
- c. 3,460 Ma – Fossils of bacteria in chert.[citation needed] Zimbabwe Craton stabilizes from the suture of two smaller crustal blocks, the Tokwe Segment to the south and the Rhodesdale Segment or Rhodesdale gneiss to the north.
- c. 3.400 Ma – Eleven taxa of prokaryotes are preserved in the Apex Chert of the Pilbara craton in Australia. Because chert is fine-grained silica-rich microcrystalline, cryptocrystalline or microfibrious material, it preserves small fossils quite well. Stabilization of Baltic Shield begins.
- c. 3.340 Ma – Johannesburg Dome forms in South Africa: located in the central part of Kaapvaal craton and consists of trondhjemitic and tonalitic granitic rocks intruded into mafic-ultramafic greenstone – the oldest granitoid phase recognised so far.
- c. 3,300 Ma – Onset of compressional tectonics. Intrusion of granitic plutons on the Kaapvaal craton.
- c. 3,260 Ma – One of the largest recorded impact events occurs near the Barberton Greenstone Belt, when a 58 km (36 mi) asteroid leaves a crater almost 480 km (300 mi) across – two and a half times larger in diameter than the Chicxulub crater.
Mesoarchean Era
- c. 3,200 Ma – Mesoarchean Era starts. Onverwacht series in South Africa form – contain some of the oldest microfossils mostly spheroidal and carbonaceous alga-like bodies.
- c. 3,200–2,600 Ma – Assembly of the Ur supercontinent to cover between 12 and 16% of the current continental crust. Formation of Limpopo Belt.
- c. 3,100 Ma – Fig Tree Formation: second round of fossilizations including Archaeosphaeroides barbertonensis and Eobacterium. Gneiss and greenstone belts in the Baltic Shield are laid down in Kola Peninsula, Karelia and northeastern Finland.
- c. 3,000 Ma – Humboldt Orogeny in Antarctica: possible formation of Humboldt Mountains in Queen Maud Land. Photosynthesizing cyanobacteria evolve; they use water as a reducing agent, thereby producing oxygen as a waste product. The oxygen initially oxidizes dissolved iron in the oceans, creating iron ore – over time oxygen concentration in the atmosphere slowly rises, acting as a poison for many bacteria. As the Moon is still very close to Earth and causes tides 1,000 feet (305 m) high[citation needed], the Earth is continually wracked by hurricane-force winds – these extreme mixing influences are thought to stimulate evolutionary processes. Rise of Stromatolites: microbial mats become successful forming the first reef building communities on Earth in shallow warm tidal pool zones (to 1.5 Gyr). Tanzania Craton forms.
- c. 2,940 Ma – Yilgarn craton of western Australia forms by the accretion of a multitude of formerly present blocks or terranes of existing continental crust.
- c. 2,900 Ma – Assembly of the Kenorland supercontinent, based upon the core of the Baltic shield, formed at c.3100 Ma. Narryer gneiss terrane (including Jack Hills) of Western Australia undergoes extensive metamorphism.
Neoarchean Era
- c. 2,800 Ma – Neoarchean Era starts. Breakup of the Vaalbara: Breakup of supercontinent Ur as it becomes a part of the major supercontinent Kenorland. Kaapvaal and Zimbabwe cratons join together.
- c. 2,770 Ma – Formation of Hamersley Basin on the southern margin of Pilbara Craton – last stable submarine-fluviatile environment between the Yilgarn and Pilbara prior to rifting, contraction and assembly of the intracratonic Gascoyne Complex.
- c. 2,750 Ma – Renosterkoppies Greenstone Belt forms on the northern edge of the Kaapvaal craton.
- c. 2,736 Ma – Formation of the Temagami Greenstone Belt in Temagami, Ontario, Canada.
- c. 2,707 Ma – Blake River Megacaldera Complex begins to form in present-day Ontario and Quebec – first known Precambrian supervolcano – first phase results in creation of 8 km long, 40 km wide, east–west striking Misema Caldera* – coalescence of at least two large mafic shield volcanoes.
- c. 2,705 Ma – Major komatiite eruption, possibly global – possible mantle overturn event.
- c. 2,704 Ma – Blake River Megacaldera Complex: second phase results in creation of 30 km long, 15 km wide northwest–southeast trending New Senator Caldera – thick massive mafic sequences which has been inferred to be a subaqueous lava lake.
- c. 2,700 Ma – Biomarkers of cyanobacteria discovered, together with steranes (sterols of cholesterol), associated with films of eukaryotes, in shales located beneath banded iron formation hematite beds, in Hamersley Range, Western Australia; skewed sulfur isotope ratios found in pyrites show a small rise in oxygen concentration in the atmosphere; Sturgeon Lake Caldera forms in Wabigoon greenstone belt – contains well preserved homoclinal chain of greenschist facies, metamorphosed intrusive, volcanic and sedimentary layers (Mattabi pyroclastic flow considered third most voluminous eruptive event); stromatolites of Bulawayo series in Zimbabwe form – first verified reef community on Earth.
- c. 2,696 Ma – Blake River Megacaldera Complex: third phase of activity constructs classic east-northeast striking Noranda Caldera which contains a 7-to-9-km-thick succession of mafic and felsic rocks erupted during five major series of activity. Abitibi greenstone belt in present-day Ontario and Quebec begins to form: considered world's largest series of Archean greenstone belts, appears to represent a series of thrusted subterranes.
- c. 2,690 Ma – Formation of high pressure granulites in the Limpopo Central Region.
- c. 2,650 Ma – Insel Orogeny: occurrence of a very high grade discrete tectonothermal event (a UHT metamorphic event).
- c. 2,600 Ma – Oldest known giant carbonate platform. Saturation of oxygen in ocean sediments is reached as oxygen now begins to dramatically appear in Earth's atmosphere.
Proterozoic Eon
Main article: Proterozoic
The Proterozoic (from c. 2500 Ma to c. 539 Ma) saw the first traces of biological activity. Fossil remains of bacteria and algae.
Paleoproterozoic Era
Main article: Paleoproterozoic
Siderian Period
- c. 2,500 Ma – Proterozoic Eon, Paleoproterozoic Era, and Siderian Period start. Oxygen saturation in the oceans is reached: Banded iron formations form and saturate ocean floor deposits – without an oxygen sink, Earth's atmosphere becomes highly oxygenic. Great Oxidation Event led by cyanobacteria's oxygenic photosynthesis – various forms of Archaea and anoxic bacteria become extinct in first great extinction event on Earth. Algoman Orogeny or Kenoran: assembly of Arctica out of the Canadian Laurentian Shield and Siberian craton – formation of Angaran Shield and Slave Province.
- c. 2,440 Ma – Formation of Gawler craton in Australia.
- c. 2,400 Ma – Huronian glaciation starts, probably from oxidation of earlier methane greenhouse gas produced by burial of organic sediments of photosynthesizers. Formation of Dharwar Craton in southern India.
- c. 2,400 Ma – Dharwar Craton in southern India stabilizes.
Rhyacian Period
- c. 2,300 Ma – Rhyacian period starts.
- c. 2,250 Ma – Bushveld Igneous Complex forms: world's largest reserves of platinum-group metals (platinum, palladium, osmium, iridium, rhodium and ruthenium), as well as vast quantities of iron, tin, chromium, titanium and vanadium appear – formation of Transvaal Basin begins.
- c. 2,200–1800 Ma – Continental Red Beds found, produced by iron in weathered sandstone being exposed to oxygen. Eburnean Orogeny, series of tectonic, metamorphic and plutonic events establish Eglab Shield to the north of West African Craton and Man Shield to its south – Birimian domain of West Africa established and structured.
- c. 2,200 Ma – Iron content of ancient fossil soils shows an oxygen built up to 5–18% of current levels. End of Kenoran Orogeny: invasion of Superior and Slave Provinces by basaltic dikes and sills – Wyoming and Montana arm of Superior Province experiences intrusion of 5 km thick sheet of chromite-bearing gabbroic rock as Stillwater Complex forms.
- c. 2,100 Ma – Huronian glaciation ends. Earliest known eukaryote fossils found. Earliest multicellular organisms collectively referred to as the "Gabonionta" (Francevillian Group Fossil); Wopmay orogeny along western margin of Canadian Shield.
- c. 2,090 Ma – Eburnean Orogeny: Eglab Shield experiences syntectonic trondhjemitic pluton intrusion of its Chegga series – most of the intrusion is in the form of a plagioclase called oligoclase.
- 2.070 Ma – Eburnean Orogeny: asthenospheric upwelling releases large volume of post-orogenic magmas – magma events repeatedly reactivated from the Neoproterozoic to the Mesozoic.
Orosirian Period
- c. 2,050 Ma – Orosirian Period starts. Significant orogeny in most continents.
- c. 2,023 Ma – Vredefort impact structure forms.
- c. 2,005 Ma – Glenburgh Orogeny (to c. 1,920 Ma) begins: Glenburgh terrane in western Australia begins to stabilize during period of substantial granite magmatism and deformation; Halfway Gneiss and Moogie Metamorphics result. Dalgaringa Supersuite (to c. 1,985 Ma), comprising sheets, dykes and viens of mesocratic and leucocratic tonalite, stabilizes.
- c. 2,000 Ma – The lesser supercontinent Atlantica forms. The Oklo natural nuclear reactor of Gabon produced by uranium-precipitant bacteria.
- c. 1,900–,880 Ma – Gunflint chert biota forms flourishes including prokaryotes like Kakabekia, Gunflintia, Animikiea and Eoastrion
- c. 1,850 Ma – Sudbury impact structure. Penokean orogeny. Bacterial viruses (bacteriophage) emerge before, or soon after, the divergence of the prokaryotic and eukaryotic lineages.
- c. 1,830 Ma – Capricorn Orogeny (1.83–1.78 Gyr) stabilizes central and northern Gascoyne Complex: formation of pelitic and psammitic schists known as Morrissey Metamorphics and depositing Pooranoo Metamorphics an amphibolite facies
Statherian Period
- c. 1,800 Ma – Statherian Period starts. Supercontinent Columbia forms, one of whose fragments being Nena. Oldest ergs develop on several cratons Barramundi Orogeny (c. 1.8 Gyr) influences MacArthur Basin in Northern Australia.
- c. 1,780 Ma – Colorado Orogeny (1.78 – 1.65 Gyr) influences southern margin of Wyoming craton–collision of Colorado orogen and Trans-Hudson orogen with stabilized Archean craton structure
- c. 1,770 Ma – Big Sky Orogeny (1.77 Gyr) influences southwest Montana: collision between Hearne and Wyoming cratons
- c. 1,765 Ma – As Kimban Orogeny in Australian continent slows, Yapungku Orogeny (1.765 Gyr) begins affecting Yilgarn craton in Western Australia – possible formation of Darling Fault, one of longest and most significant in Australia
- c. 1,760 Ma – Yavapai Orogeny (1.76–1.7 Gyr) impacts mid- to south-western United States; Concentrated uranium deposits in Oklo, Gabon, in West Africa are activated after being inundated with ground water in what amounts to a natural nuclear reaction – Reactions continue off and on probably never exceeding 100 kilowatts of thermal power during this time
- c. 1,750 Ma – Gothian Orogeny (1.75–1.5 Gyr): formation of tonalitic-granodioritic plutonic rocks and calc-alkaline volcanites in the East European Craton
- c. 1,700 Ma – Stabilization of second major continental mass, the Guiana Shield in South America.
- c. 1,680 Ma – Mangaroon Orogeny (1.68–1.62 Gyr), on the Gascoyne Complex in Western Australia: Durlacher Supersuite, granite intrusion featuring a northern (Minnie Creek) and southern belt – heavily sheared orthoclase porphyroclastic granites
- c. 1,650 Ma – Kararan Orogeny (1.65 Gyr) uplifts great mountains on the Gawler craton in Southern Australia – formation of Gawler Range including picturesque Conical Hill Track and "Organ Pipes" waterfall
Mesoproterozoic Era
Main article: Mesoproterozoic
Calymmian Period
- c. 1,600 Ma – Mesoproterozoic Era and Calymmian Period start. Platform covers expand. Major orogenic event in Australia: Isan Orogeny influences Mount Isa Block of Queensland – major deposits of lead, silver, copper and zinc are laid down. Mazatzal Orogeny (to c. 1,300 Ma) influences mid- to south-western United States: Precambrian rocks of the Grand Canyon, Vishnu Schist and Grand Canyon Series, are formed establishing basement of Canyon with metamorphosed gneisses that are intruded by granites. Belt Supergroup in Montana/Idaho/BC formed in basin on edge of Laurentia.
- c. 1,500 Ma – Supercontinent Columbia splits apart: associated with continental rifting along western margin of Laurentia, eastern India, southern Baltica, southeastern Siberia, northwestern South Africa and North China Block-formation of Ghats Province in India. First structurally complex eukaryotes (Horodyskia, colonial formamiferian?).
Ectasian Period
- c. 1,400 Ma – Ectasian Period starts. Platform covers expand. Major increase in Stromatolite diversity with widespread blue-green algae colonies and reefs dominating tidal zones of oceans and seas
- c. 1,300 Ma – Break-up of Columbia Supercontinent completed: widespread anorogenic magmatic activity, forming anorthosite-mangerite-charnockite-granite suites in North America, Baltica, Amazonia and North China – stabilization of Amazonian Craton in South America Grenville orogeny(to c. 1,000 Ma) in North America: globally associated with assembly of Supercontinent Rodinia establishes Grenville Province in Eastern North America – folded mountains from Newfoundland to North Carolina as Old Rag Mountain forms
- c. 1,270 Ma – Emplacement of Mackenzie granite mafic dike swarm – one of three dozen dike swarms, forms into Mackenzie Large Igneous Province – formation of Copper Creek deposits
- c. 1,250 Ma – Sveconorwegian Orogeny (to c. 900 Ma) begins: essentially a reworking of previously formed crust on the Baltic Shield
- c. 1,240 Ma – Second major dike swarm, Sudbury dikes form in Northeastern Ontario around the area of the Sudbury Basin
Stenian Period
- c. 1,200 Ma – Stenian Period starts. Red alga Bangiomorpha pubescens, earliest fossil evidence for sexually reproducing organism. Meiosis and sexual reproduction are present in single-celled eukaryotes, and possibly in the common ancestor of all eukaryotes. Supercontinent of Rodinia (1.2 Gyr–750 Ma) completed: consisting of North American, East European, Amazonian, West African, Eastern Antarctica, Australia and China blocks, largest global system yet formed – surrounded by superocean Mirovia
- c. 1,100 Ma – First dinoflagellate evolve; photosynthetic, some develop mixotrophic habits of ingesting prey. Thus, they become the first predators, forcing acritarchs to defensive strategies and leading to open "arms" race. Late Ruker (1.1–1 Gyr) and Nimrod Orogenies (1.1 Gyr) in Antarctica possibly begins: formation of Gamburtsev mountain range and Vostok Subglacial Highlands. Keweenawan Rift buckles in the south-central part of the North American plate – leaves behind thick layers of rock that are exposed in Wisconsin, Minnesota, Iowa and Nebraska and creates rift valley where future Lake Superior develops.
- c. 1,080 Ma – Musgrave Orogeny (c. 1.080 Gyr) forms Musgrave Block, an east–west trending belt of granulite-gneiss basement rocks – voluminous Kulgera Suite of granite and Birksgate Complex solidify
- c. 1,076 Ma – Musgrave Orogeny: Warakurna large igneous province develops – intrusion of Giles Complex and Winburn Suite of granites and deposition of Bentley Supergroup (including Tollu and Smoke Hill Volcanics)
- c. 1,010 Ma – Ourasphaira giraldae: multicellular organic-walled microfossils preserved in shale of the Grassy Bay Formation (Canadian Arctic) with fungal affinity.
Neoproterozoic Era
Main article: Neoproterozoic
Tonian Period
- c. 1,000 Ma – Neoproterozoic Era and Tonian Period start. Grenville orogeny ends. First radiation of dinoflagellates and spiny acritarchs – increase in defensive systems indicate that acritarchs are responding to carnivorous habits of dinoflagellates – decline in stromatolite reef populations begins. Rodinia starts to break up. First vaucherian algae. Rayner Orogeny as proto-India and Antarctica collide (to c. 900 Ma). Trace fossils of colonial Horodyskia (to c. 900 Ma): possible divergence between animal and plant kingdoms begins. Stabilization of Satpura Province in Northern India.
- c. 920 Ma – Edmundian Orogeny (c. 920–850 Ma) redefines Gascoyne Complex: consists of reactivation of earlier formed faults in the Gascoyne – folding and faulting of overlying Edmund and Collier basins
- c. 920 Ma – Adelaide Geosyncline laid down in central Australia – essentially a rift complex, consists of thick layer of sedimentary rock and minor volcanics deposited on Easter margin – limestones, shales and sandstones predominate
- c. 900 Ma – Bitter Springs Formation of Australia: in addition to prokaryote assemblage of fossils, cherts include eukaryotes with ghostly internal structures similar to green algae – first appearance of Glenobotrydion (900–720 Ma), among earliest plants on Earth.
- c. 850 Ma - Start of the Neoproterozoic oxygenation event, a second major increase in atmospheric and oceanic oxygen concentration on Earth.
- c. 830 Ma – Rift develops on Rodinia between continental masses of Australia, eastern Antarctica, India, Congo and Kalahari on one side and Laurentia, Baltica, Amazonia, West African and Rio de la Plata cratons on other – formation of Adamastor Ocean.
- c. 800 Ma – With free oxygen levels much higher, carbon cycle is disrupted and once again glaciation becomes severe – beginning of second "snowball Earth" event
- c. 750 Ma – First Protozoa appears: as creatures like Paramecium, Amoeba and Melanocyrillium evolve, first animal-like cells become distinctive from plants – rise of herbivores (plant feeders) in the food chain. First Sponge-like animal: similar to early colonial foraminiferan Horodyskia, earliest ancestors of Sponges were colonial cells that circulated food sources using flagella to their gullet to be digested. Kaigas (c. 750 Ma): first thought to be a major glaciation of Earth, however, the Kaigas formation was later determined to be non-glacial.
Cryogenian Period
- c. 720 Ma – Cryogenian Period starts, during which Earth freezes over (Snowball Earth or Slushball Earth) at least 3 times. The Sturtian glaciation continues the process begun during Kaigas – great ice sheets cover most of the planet stunting evolutionary development of animal and plant life – survival based on small pockets of heat under the ice.
- c. 700 Ma – Fossils of testate Amoeba first appear: first complex metazoans leave unconfirmed biomarkers – they introduce new complex body plan architecture which allows for development of complex internal and external structures. Worm trail impressions in China: because putative "burrows" under stromatolite mounds are of uneven width and tapering makes biological origin difficult to defend – structures imply simple feeding behaviours. Rifting of Rodinia is completed: formation of new superocean of Panthalassa as previous Mirovia ocean bed closes – Mozambique mobile belt develops as a suture between plates on Congo-Tanzania craton
- c. 660 Ma – As Sturtian glaciers retreat, Cadomian orogeny (660–540 Ma) begins on north coast of Armorica: involving one or more collisions of island arcs on margin of future Gondwana, terranes of Avalonia, Armorica and Iberia are laid down
- c. 650 Ma – First Demosponges appear: form first skeletons of spicules made from protein spongin and silica – brightly coloured these colonial creatures filter feed since they lack nervous, digestive or circulatory systems and reproduce both sexually and asexually
- c. 650 Ma – Final period of worldwide glaciation, Marinoan (650–635 Ma) begins: most significant "snowball Earth" event, global in scope and longer – evidence from Diamictite deposits in South Australia laid down on Adelaide Geosyncline
Ediacaran Period
- c. 635 Ma – Ediacaran period begins. End of Marinoan Glaciation: last major "snowball Earth" event as future ice ages will feature less overall ice coverage of the planet
- c. 633 Ma – Beardmore Orogeny (to c. 620 Ma) in Antarctica: reflection of final break-up of Rodinia as pieces of the supercontinent begin moving together again to form Pannotia
- c. 620 Ma – Timanide Orogeny (to c. 550 Ma) affects northern Baltic Shield: gneiss province divided into several north–south trending segments experiences numerous metasedimentary and metavolcanic deposits – last major orogenic event of Precambrian
- c. 602 Ma - Lantian Biota appears, which includes some of the oldest large and complex fossils known.
- c. 600 Ma – Pan-African Orogeny begins: Arabian-Nubian Shield formed between plates separating supercontinent fragments Gondwana and Pannotia – Supercontinent Pannotia (to c. 500 Ma) completed, bordered by Iapetus and Panthalassa oceans. Accumulation of atmospheric oxygen allows for the formation of ozone layer: prior to this, land-based life would probably have required other chemicals to attenuate ultraviolet radiation enough to permit colonization of the land
- c. 575 Ma – First Ediacaran-type fossils.
- c. 565 Ma – Charnia, a frond-like organism, first evolves.
- c. 560 Ma – Trace fossils, e.g., worm burrows, and small bilaterally symmetrical animals. Earliest arthropods. Earliest fungi.
- c. 558 Ma – Dickinsonia, a large slow moving disc-like creature, first appears – the discovery of fat molecules in its tissues make it the first confirmed true metazoan animal of the fossil record.
- c. 555 Ma – The first possible mollusk Kimberella appears.
- c. 550 Ma – First possible comb-jellies, sponges, corals, and anemones.
- c. 550 Ma – Uluru or Ayers Rock begins forming during the Petermann Orogeny in Australia
- c. 544 Ma – The small shelly fauna first appears.
Phanerozoic Eon
Main article: Phanerozoic
Paleozoic Era
Main article: Paleozoic
Cambrian Period
- c. 538.8 ± 0.6 Ma – beginning of the Cambrian Period, the Paleozoic Era and the current Phanerozoic Eon. End of the Ediacaran Period, the Proterozoic Eon and the Precambrian Supereon. The Ediacaran fauna disappears, while the Cambrian explosion initiates the emergence of most forms of complex life, including vertebrates (fish), arthropods, echinoderms and molluscs. Pannotia breaks up into several smaller continents: Laurentia, Baltica and Gondwana.
- c. 530 Ma – First fish – appearance of Myllokunmingia
- c. 525 Ma – First graptolites.
- c. 521 Ma – First trilobites.
- c. 518 Ma – Chengjiang biota flourishes – Maotianshan Shales reveal numerous invertebrates and arthropods that appear in the Burgess shales suggesting their range is global and includes a number of chordates including Haikouella, Yunnanozoon and early fish like Haikouichthys.
- c. 514 Ma – Paradoxides trilobites appear, the largest members of the Cambrian Trilobites.
- c. 511 Ma – Earliest crustaceans.
- c. 505 Ma – Deposition of the Burgess Shale – Biota includes numerous strange invertebrates and arthropods like Opabinia; First great apex predator Anomalocaris dominates.
- c. 490 Ma – Beginning of the Caledonian Orogeny as three continents and terranes of Laurentia, Baltica and Avalonia collide resulting in mountain-building recorded in the northern parts of Ireland and Britain, the Scandinavian Mountains, Svalbard, eastern Greenland and parts of north-central Europe.
- c. 488 Ma – Earliest brittle stars.
Ordovician Period
- c. 486.85 ± 1.5 Ma – Beginning of the Ordovician and the end of the Cambrian Period.
- c. 485 Ma – First jawless fish – radiation of Thelodont fish into the Silurian
- c. 460 Ma – First crinoids evolve.
- c. 450 Ma – Late Ordovician microfossils of scales indicate the earliest evidence for the existence of jawed fish or Gnathostomata.
- c. 450 Ma – Plants and arthropods colonize the land. Sharks evolve. First horseshoe crabs and starfish.
Silurian Period
- c. 443.1 ± 0.9 Ma – Beginning of the Silurian and the end of the Ordovician Period.
- c. 433 Ma – Great Glen Fault begins shaping the Scottish Highlands as the Caledonian Orogeny reaches its close.
- c. 430 Ma – First appearance of Cooksonia, the oldest known plant to have a stem with vascular tissue and is thus a transitional form between the primitive non-vascular bryophytes and the vascular plants
- c. 420 Ma – First creature took a breath of air. First ray-finned fish and land scorpions.
Devonian Period
- c. 419.62 ± 1.36 Ma – Beginning of the Devonian and end of the Silurian Period. First insects.
- c. 419 Ma – Old Red Sandstone sediments begin being laid in the North Atlantic region including Britain, Ireland, Norway and in the west along the northeastern seaboard of North America. It also extends northwards into Greenland and Svalbard.
- c. 415 Ma – Cephalaspis, an iconic member of the Osteostraci, appears, the most advanced of the jawless fish. Its boney armor serves as protection against the successful radiation of Placoderms and as a way to live in calcium-poor fresh water environments.
- c. 410 Ma – First toothed fish and nautiloids.
- c. 395 Ma – First of many modern groups, including tetrapods.
- c. 375 Ma – Acadian Orogeny begins influencing mountain building along the Atlantic seaboard of North America.
- c. 370 Ma – Cladoselache, an early shark, first appears.
- c. 363 Ma – Vascular plants begin to create the earliest stable soils on land.
- c. 360 Ma – First crabs and ferns. The large predatory lobe-finned fish Hyneria evolves.
Carboniferous Period
- c. 358.86 ± 0.14 Ma – Beginning of the Carboniferous and the end of Devonian Period. Amphibians diversify. Coal forests appear.
- c. 350 Ma – First large sharks, ratfish and hagfish.
- c. 345 Ma – Agaricocrinus americanus a representative of the Crinoids appears as part of a successful radiation of the echinoderms.
- c. 330 Ma – First amniotes evolve.
- c. 320 Ma – First sauropsids evolve.
- c. 318 Ma – First synapsids evolve.
- c. 312 Ma – Hylonomus makes first appearance, one of the oldest reptiles found in the fossil record.
- c. 306 Ma – Diplocaulus evolves in the swamps with an unusual boomerang-like skull.
- c. 305 Ma – Carboniferous rainforest collapse. First diapsids evolve. Meganeura, a giant dragonfly, dominates the skies.
- c. 300 Ma – Last great period of mountain building episodes in Europe and North America in response to the final suturing together of the supercontinent Pangaea – the Ural Mountains are uplifted
Permian Period
- c. 298.9 ± 0.15 Ma – End of Carboniferous and beginning of Permian Period. By this time, all continents have fused into the supercontinent of Pangaea. Seed plants and conifers diversify along with temnospondyls and pelycosaurs.
- c. 296 Ma – Oldest known octopus fossil.
- c. 295 Ma – Dimetrodon evolves.
- c. 295 Ma – First beetles appear.
- c. 280 Ma – First cycads evolve.
- c. 275 Ma – First therapsids evolve.
- c. 270 Ma – Gorgonopsians, the apex predators of the Late Permian, first evolve.
- c. 251.9 Ma – Permian mass extinction. End of Permian Period and of the Palaeozoic Era. Beginning of Triassic Period, the Mesozoic era and of the age of the dinosaurs.
Mesozoic Era
Main article: Mesozoic
Triassic Period
- c. 251.9 Ma ± 0.024 Ma – Mesozoic era and Triassic Period begin. Mesozoic Marine Revolution begins.
- c. 247 Ma - First sauropterygians.
- c. 245 Ma – First ichthyosaurs.
- c. 240 Ma – Cynodonts and rhynchosaurs diversify.
- c. 233 Ma — Earliest sauropods appear.
- c. 231 Ma - First theropods appear.
- c. 228 Ma - Pterosaurs evolve.
- c. 227 Ma - First plesiosaurs.
- c. 225 Ma – First teleosts evolve. Also the earliest appearance of mammals.
- c. 220 Ma – First crocodilians and flies.
- c. 215 Ma – First turtles. Coelophysis, one of the earliest known dinosaurs, appears.
- c. 214 Ma - Plateosaurus, a basal sauropodomorph or so-called "prosauropod" evolves in what is now Central and Northern Europe, Greenland and North America
- c. 210 Ma – Earliest elasmosauridae.
Jurassic Period
- c. 201.4 ± 0.2 Ma – Triassic-Jurassic extinction event marks the end of Triassic and beginning of Jurassic Period. The largest dinosaurs, such as Diplodocus and Brachiosaurus evolve during this time, as do the carnosaurs; large, bipedal predatory dinosaurs such as Allosaurus. First specialized pterosaurs and sauropods. Ornithischians diversify.
- c. 199 Ma – First squamata evolve. Earliest lizards.
- c. 190 Ma – Pliosaurs evolve, along with many groups of primitive sea invertebrates.
- c. 190 Ma - Thalattosuchians evolve.
- c. 180 Ma – Pangaea splits into two major continents: Laurasia in the north and Gondwana in the south.
- c. 176 Ma – First stegosaurs.
- c. 170 Ma – First salamanders and newts evolve. First megalosaurs. Cynodonts go extinct.
- c. 167 Ma - First crown group mammals.
- c. 165 Ma – First rays and glycymeridid bivalves.
- c. 164 Ma – The first gliding mammal, volaticotherium, appears in the fossil record.
- c. 161 Ma – First ceratopsians.
- c. 155 Ma – First birds and triconodonts. Stegosaurs and theropods diversify.
- c. 153 Ma – Earliest pine trees.
- c. 145 Ma – First mantises.
Cretaceous Period
- c. 143.1 ± 0.6 Ma – End of Jurassic and beginning of Cretaceous Period. First dromeosaurs (raptors) evolve.
- c. 140 Ma – Earliest orb-weaver spiders evolve. First titanosaurs.
- c. 130 Ma – Laurasia and Gondwana begin to split apart as the Atlantic Ocean forms. First flowering plants. Earliest anglerfish.
- c. 125 Ma – Sinodelphys szalayi, the earliest known marsupial, evolves in China.
- c. 122 Ma – Earliest ankylosauridae.
- c. 115 Ma – First monotremes.
- c. 110 Ma – First hesperornithes.
- c. 106 Ma – Spinosaurus evolves.
- c. 100.5 Ma - Stegosaurs go extinct
- c. 100 Ma – First bees.
- c. 94 Ma - First mosasaurs.
- c. 93 Ma - Cenomanian-Turonian boundary event causes the extinction of ichthyosaurs and pliosaurs.
- c. 90 Ma – the Indian subcontinent splits from Gondwana, becoming an island continent. Snakes and ticks evolve. Carnosaurs (comprising megalosaurids, allosaurids and carcharodontosaurids) go extinct.
- c. 86 Ma – First hadrosauridae.
- c. 80 Ma – Australia splits from Antarctica. First ants. First palm trees appear.
- c. 75 Ma – First velociraptors.
- c. 70 Ma – Multituberculates diversify. The Mosasaurus evolves.
- c. 68 Ma – Tyrannosaurus rex evolves. Earliest species of Triceratops. Quetzalcoatlus, one of the largest flying animals to ever live, first appears in the fossil record.
- c. 66.038 ± 0.011 Ma – Cretaceous-Paleogene extinction event at the end of the Cretaceous Period marks the end of the Mesozoic era and the age of the dinosaurs; start of the Paleogene Period and the current Cenozoic era.
Cenozoic Era
Main article: Cenozoic
Paleogene Period
- c. 63 Ma – First creodonts.
- c. 62 Ma – Fall in sea level and the retreat of inland seas completes the emergence of North America; First penguins appear – Genus Crossvallia, the earliest known birds suited to an aquatic lifestyle, alongside Kupoupou appear in the fossil record of Antarctica (from the Cross Valley Formation on Seymour Island) and New Zealand (Takatika Grit formation of the Chatham Islands); Pelagornithidae or bony-toothed birds, group of large seabirds noted for tooth-like points on their beak's edges first appear – Pseudoteeth of pelagornithids do not seem to have had serrated or otherwise specialized cutting edges and were useful to hold prey for swallowing whole and would have consisted of soft-bodied organisms like Cephalopods; .
- c. 60 Ma – Evolution of the first primates and miacids. Flightless birds diversify.
- c. 56 Ma – Gastornis evolves.
- c. 55 Ma – the island of the Indian subcontinent collides with Asia, thrusting up the Himalayas and the Tibetan Plateau. Many modern bird groups appear. First whale ancestors. First rodents, lagomorphs, armadillos, sirenians, proboscideans, perissodactyls, artiodactyls, and mako sharks. Angiosperms diversify.
- c. 52.5 Ma – First passerine (perching) birds.
- c. 52 Ma – First bats.
- c. 50 Ma – Africa collides with Eurasia, closing the Tethys Sea. Divergence of cat and dog ancestors. Primates diversify. Brontotheres, tapirs, and rhinos evolve.
- c. 49 Ma – Whales return to the water.
- c. 45 Ma – Camels evolve in North America.
- c. 40 Ma – Age of the Catarrhini parvorder; first canines evolve. Lepidopteran insects become recognizable. Gastornis goes extinct. Basilosaurus evolves.
- c. 37 Ma – First Nimravids.
- c. 33.9 Ma – End of Eocene, start of Oligocene epoch.
- c. 35 Ma – Grasslands first appear. Glyptodonts, ground sloths, peccaries, dogs, eagles, and hawks evolve.
- c. 33 Ma – First thylacinid marsupials evolve.
- c. 30 Ma – Brontotheres go extinct. Pigs evolve. South America separates from Antarctica, becoming an island continent.
- c. 28 Ma – Paraceratherium evolves. First pelicans.
- c. 26 Ma – Emergence of the first true elephants.
- c. 25 Ma – First deers. Felids evolve.
- c. 24 Ma – Earliest pinnipeds (seals).
Neogene Period
|
Hominin timeline
|
||||||||||||||||||||||||||||||
|
−10 —
–
−9 —
–
−8 —
–
−7 —
–
−6 —
–
−5 —
–
−4 —
–
−3 —
–
−2 —
–
−1 —
–
0 —
|
Miocene
Pliocene
Pleistocene
Hominini
Nakalipithecus
Samburupithecus
Ouranopithecus
(Ou. turkae) (Ou. macedoniensis) Chororapithecus
Oreopithecus
Sivapithecus
Sahelanthropus
Graecopithecus
Orrorin
(O. praegens)
(O. tugenensis) Ardipithecus
(Ar. kadabba)
(Ar. ramidus)
Australopithecus
(Au. africanus) (Au. afarensis) (Au. anamensis) H. habilis
(H. rudolfensis) (Au. garhi) H. erectus
(H. antecessor) (H. ergaster) (Au. sediba) H. heidelbergensis
Homo sapiens
Neanderthals
Denisovans
|
|
||||||||||||||||||||||||||||
|
(million years ago)
|
||||||||||||||||||||||||||||||
- c. 23.04 Ma – Neogene Period and Miocene epoch begin
- c. 22 Ma – First hyenas.
- c. 20 Ma – Giraffes and giant anteaters evolve.
- c. 18–12 Ma – estimated age of the Hominidae/Hylobatidae (great apes vs. gibbons) split.
- c. 16 Ma – The hippopotamus evolves.
- c. 15 Ma – First mastodons, bovids, and kangaroos. Australian megafauna diversify.
- c. 11 Ma – Estimated date for the origin of the modern Yangtze river.
- c. 10 Ma – Insects diversify. First large horses. Camels cross from America to Asia.
- c. 6.5 Ma – First members of the Hominini tribe.
- c. 6 Ma – Australopithecines diversify.
- c. 5.96–5.33 Ma – Messinian Salinity Crisis: the precursor of the current Strait of Gibraltar closes repeatedly, leading to a partial desiccation and strong increase in salinity of the Mediterranean Sea.
- c. 5.4–6.3 Ma – Estimated age of the Homo/Pan (human vs. chimpanzee) split.
- c. 5.5 Ma – Appearance of the genus Ardipithecus
- c. 5.33 Ma – Zanclean flood: the Strait of Gibraltar opens for the last (and current) time and water from the Atlantic Sea fills again the Mediterranean Sea basin. The deep canyon carved by the Eonile during the Messinian Salinity Crisis is filled with seawater up to at least Aswan. The modern Nile starts filling this sea branch with sediments, slowly creating the Nile Valley.
- c. 5.333 Ma – Pliocene epoch begins. First tree sloths. First large vultures. Nimravids go extinct.
- c. 5.0 Ma – The Colorado Plateau reaches its present height, and the course of the Colorado River becomes close to the present one.
- c. 4.8 Ma – The mammoth appears.
- c. 4.2 Ma – appearance of the genus Australopithecus
- c. 4 Ma – First zebras.
- c. 3 Ma – Isthmus of Panama joins North and South America. Great American Interchange. Cats, condors, raccoons and camelids move south; armadillos, hummingbirds, and opossums move north.
- c. 2.7 Ma – Paranthropus evolves.
- c. 2.6 Ma – The current ice age begins.
Quaternary Period
- c. 2.58 Ma – start of the Pleistocene epoch, the Stone Age and the current Quaternary Period; emergence of the genus Homo. Smilodon, the best known of the sabre-toothed cats, appears.
- c. 2.4 Ma – The Amazon River takes its present shape in South America.
- c. 2.0–1.5 Ma – The basin of the Congo River acquires its present shape.
- c. 1.9 Ma – Oldest known Homo erectus fossils. This species might be evolved some time before, up to c. 2 Ma ago.
- c. 1.7 Ma – Australopithecines go extinct.
- c. 1.8–0.8 Ma – colonisation of Eurasia by Homo erectus.
- c. 1.5 Ma – earliest possible evidence of the controlled use of fire by Homo erectus
- c. 1.2 Ma – Homo antecessor evolves. Paranthropus dies out.
- c. 0.79 Ma – earliest demonstrable evidence of the controlled use of fire by Homo erectus
- c. 0.7 Ma – last reversal of the Earth's magnetic field
- c. 0.7 Ma: oldest archaic hominins that broke away from the modern human lineage that were found to have inserted into the Sub-Saharan African population genome approximately 35,000 years ago.
- c. 0.64 Ma – Yellowstone caldera erupts
- c. 0.6 Ma – Homo heidelbergensis evolves.
- c. 0.5 Ma – First brown bears.
- c. 0.315 Ma – Middle Paleolithic begins. Appearance of Homo sapiens in Africa
Hadean Eon

Main article: Hadean
4540 Ma – 4031 Ma
| Date | Event |
|---|---|
| 4540 Ma | Planet Earth forms from the accretion disc revolving around the young Sun, perhaps preceded by formation of organic compounds necessary for life in the surrounding protoplanetary disk of cosmic dust. |
| 4510 Ma | According to the giant-impact hypothesis, the Moon originated when Earth and the hypothesized planet Theia collided, sending into orbit myriad moonlets which eventually coalesced into our single Moon. The Moon's gravitational pull stabilised Earth's fluctuating axis of rotation, setting up regular climatic conditions favoring abiogenesis. |
| 4404 Ma | Evidence of the first liquid water on Earth which were found in the oldest known zircon crystals. |
| 4280–3770 Ma | Earliest possible appearance of life on Earth. |
Archean Eon
Main article: Archean


4031 Ma – 2500 Ma
| Date | Event |
|---|---|
| 4100 Ma | Earliest possible preservation of biogenic carbon. |
| 4100–3800 Ma | Late Heavy Bombardment (LHB): extended barrage by meteoroids impacting the inner planets. Thermal flux from widespread hydrothermal activity during the LHB may have aided abiogenesis and life's early diversification. Possible remains of biotic life were found in 4.1 billion-year-old rocks in Western Australia. |
| 4000 Ma | Formation of a greenstone belt of the Acasta Gneiss of the Slave craton in northwest Canada - the oldest known rock belt. |
| 3900–2500 Ma | Cells resembling prokaryotes appear. These first organisms are believed to have been chemoautotrophs, using carbon dioxide as a carbon source and oxidizing inorganic materials to extract energy. |
| 3800 Ma | Formation of a greenstone belt of the Isua complex in western Greenland, whose isotope frequencies suggest the presence of life. The earliest evidence for life on Earth includes: 3.8 billion-year-old biogenic hematite in a banded iron formation of the Nuvvuagittuq Greenstone Belt in Canada; graphite in 3.7 billion-year-old metasedimentary rocks in western Greenland; and microbial mat fossils in 3.48 billion-year-old sandstone in Western Australia. |
| 3800–3500 Ma | Last universal common ancestor (LUCA): split between bacteria and archaea. Bacteria develop primitive photosynthesis, which at first did not produce oxygen. These organisms exploit a proton gradient to generate adenosine triphosphate (ATP), a mechanism used by virtually all subsequent organisms. |
| 3000 Ma | Photosynthesizing cyanobacteria using water as a reducing agent and producing oxygen as a waste product. Free oxygen initially oxidizes dissolved iron in the oceans, creating iron ore. Oxygen concentration in the atmosphere slowly rises, poisoning many bacteria and eventually triggering the Great Oxygenation Event. |
| 2800 Ma | Oldest evidence for microbial life on land in the form of organic matter-rich paleosols, ephemeral ponds and alluvial sequences, some bearing microfossils. |
Proterozoic Eon
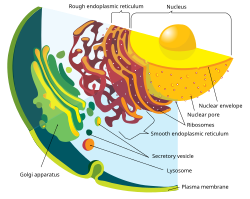

Main article: Proterozoic
2500 Ma – 539 Ma. Contains the Palaeoproterozoic, Mesoproterozoic and Neoproterozoic eras.
| Date | Event |
|---|---|
| 2500 Ma | Great Oxidation Event led by cyanobacteria's oxygenic photosynthesis. Commencement of plate tectonics with old marine crust dense enough to subduct. |
| 2023 Ma | Formation of the Vredefort impact structure, one of the largest and oldest verified impact structures on Earth. The crater is estimated to have been between 170–300 kilometres (110–190 mi) across when it first formed. |
| By 1850 Ma | Eukaryotic cells, containing membrane-bound organelles with diverse functions, probably derived from prokaryotes engulfing each other via phagocytosis. (See Symbiogenesis and Endosymbiont). Bacterial viruses (bacteriophages) emerge before or soon after the divergence of the prokaryotic and eukaryotic lineages. Red beds show an oxidising atmosphere, favouring the spread of eukaryotic life. |
| 1500 Ma | Volyn biota, a collection of exceptionally well-preserved microfossils with varying morphologies. |
| 1300 Ma | Earliest land fungi. |
| By 1200 Ma | Meiosis and sexual reproduction in single-celled eukaryotes, possibly even in the common ancestor of all eukaryotes or in the RNA world. Sexual reproduction may have increased the rate of evolution. |
| By 1000 Ma | First non-marine eukaryotes move onto land. They were photosynthetic and multicellular, indicating that plants evolved much earlier than originally thought. |
| 750 Ma | Beginning of animal evolution. |
| 720–630 Ma | Possible global glaciation which increased the atmospheric oxygen and decreased carbon dioxide, and was either caused by land plant evolution or resulted in it. Opinion is divided on whether it increased or decreased biodiversity or the rate of evolution. |
| 600 Ma | Accumulation of atmospheric oxygen allows the formation of an ozone layer. Previous land-based life would probably have required other chemicals to attenuate ultraviolet radiation. |
| 580–542 Ma | Ediacaran biota, the first large, complex aquatic multicellular organisms. |
| 580–500 Ma | Cambrian explosion: most modern animal phyla appear. |
| 550–540 Ma | Ctenophora (comb jellies), Porifera (sponges), Anthozoa (corals and sea anemones), Ikaria wariootia (an early Bilaterian). |
Phanerozoic Eon
Main article: Phanerozoic
539 Ma – present
The Phanerozoic Eon (Greek: period of well-displayed life) marks the appearance in the fossil record of abundant, shell-forming and/or trace-making organisms. It is subdivided into three eras, the Paleozoic, Mesozoic and Cenozoic, with major mass extinctions at division points.
Palaeozoic Era
Main article: Paleozoic
|
|
This section needs additional citations for verification. Please help improve this article by adding citations to reliable sources in this section. Unsourced material may be challenged and removed.
Find sources: "Timeline of the evolutionary history of life" – news · newspapers · books · scholar · JSTOR (September 2022) (Learn how and when to remove this message) |
538.8 Ma – 251.9 Ma and contains the Cambrian, Ordovician, Silurian, Devonian, Carboniferous and Permian periods.




| Date | Event |
|---|---|
| 535 Ma | Major diversification of living things in the oceans: arthropods (e.g. trilobites, crustaceans), chordates, echinoderms, molluscs, brachiopods, foraminifers and radiolarians, etc. |
| 530 Ma | The first known footprints on land date to 530 Ma. |
| 520 Ma | Earliest graptolites. |
| 511 Ma | Earliest crustaceans. |
| 505 Ma | Fossilization of the Burgess Shale |
| 500 Ma | Jellyfish have existed since at least this time. |
| 485 Ma | First vertebrates with true bones (jawless fishes). |
| 450 Ma | First complete conodonts and echinoids appear. |
| 440 Ma | First agnathan fishes: Heterostraci, Galeaspida, and Pituriaspida. |
| 420 Ma | Earliest ray-finned fishes, trigonotarbid arachnids, and land scorpions. |
| 410 Ma | First signs of teeth in fish. Earliest Nautilida, lycophytes, and trimerophytes. |
| 488–400 Ma | First cephalopods (nautiloids) and chitons. |
| 395 Ma | First lichens, stoneworts. Earliest harvestmen, mites, hexapods (springtails) and ammonoids. The earliest known tracks on land named the Zachelmie trackways which are possibly related to icthyostegalians. |
| 375 Ma | Tiktaalik, a lobe-finned fish with some anatomical features similar to early tetrapods. It has been suggested to be a transitional species between fish and tetrapods. |
| 365 Ma | Acanthostega is one of the earliest vertebrates capable of walking. |
| 363 Ma | By the start of the Carboniferous Period, the Earth begins to resemble its present state. Insects roamed the land and would soon take to the skies; sharks swam the oceans as top predators, and vegetation covered the land, with seed-bearing plants and forests soon to flourish. Four-limbed tetrapods gradually gain adaptations which will help them occupy a terrestrial life-habit. |
| 360 Ma | First crabs and ferns. Land flora dominated by seed ferns. The Xinhang forest grows around this time. |
| 350 Ma | First large sharks, ratfishes, and hagfish; first crown tetrapods (with five digits and no fins and scales). |
| 350 Ma | Diversification of amphibians. |
| 325-335 Ma | First Reptiliomorpha. |
| 330-320 Ma | First amniote vertebrates (Paleothyris). |
| 320 Ma | Synapsids (precursors to mammals) separate from sauropsids (reptiles) in late Carboniferous. |
| 305 Ma | The Carboniferous rainforest collapse occurs, causing a minor extinction event, as well as paving the way for amniotes to become dominant over amphibians and seed plants over ferns and lycophytes. First diapsid reptiles (e.g. Petrolacosaurus). |
| 280 Ma | Earliest beetles, seed plants and conifers diversify while lepidodendrids and sphenopsids decrease. Terrestrial temnospondyl amphibians and pelycosaurs (e.g. Dimetrodon) diversify in species. |
| 275 Ma | Therapsid synapsids separate from pelycosaur synapsids. |
| 265 Ma | Gorgonopsians appear in the fossil record. |
| 251.9–251.4 Ma | The Permian–Triassic extinction event eliminates over 90-95% of marine species. Terrestrial organisms were not as seriously affected as the marine biota. This "clearing of the slate" may have led to an ensuing diversification, but life on land took 30 million years to completely recover. |
Mesozoic Era
Main article: Mesozoic
|
|
This section needs additional citations for verification. Please help improve this article by adding citations to reliable sources in this section. Unsourced material may be challenged and removed.
Find sources: "Timeline of the evolutionary history of life" – news · newspapers · books · scholar · JSTOR (September 2022) (Learn how and when to remove this message) |




From 251.9 Ma to 66 Ma and containing the Triassic, Jurassic and Cretaceous periods.
| Date | Event |
|---|---|
| 250 Ma | Mesozoic marine revolution begins: increasingly well adapted and diverse predators stress sessile marine groups; the "balance of power" in the oceans shifts dramatically as some groups of prey adapt more rapidly and effectively than others. |
| 250 Ma | Triadobatrachus massinoti is the earliest known frog. |
| 248 Ma | Sturgeon and paddlefish (Acipenseridae) first appear. |
| 245 Ma | Earliest ichthyosaurs |
| 240 Ma | Increase in diversity of cynodonts and rhynchosaurs |
| 225 Ma | Earliest dinosaurs (prosauropods), first cardiid bivalves, diversity in cycads, bennettitaleans, and conifers. First teleost fishes. First mammals (Adelobasileus). |
| 220 Ma | Seed-producing Gymnosperm forests dominate the land; herbivores grow to huge sizes to accommodate the large guts necessary to digest the nutrient-poor plants.[citation needed] First flies and turtles (Odontochelys). First coelophysoid dinosaurs. First mammals from small-sized cynodonts, which transitioned towards a nocturnal, insectivorous, and endothermic lifestyle. |
| 205 Ma | Massive Triassic/Jurassic extinction. It wipes out all pseudosuchians except crocodylomorphs, who transitioned to an aquatic habitat, while dinosaurs took over the land and pterosaurs filled the air. |
| 200 Ma | First accepted evidence for viruses infecting eukaryotic cells (the group Geminiviridae). However, viruses are still poorly understood and may have arisen before "life" itself, or may be a more recent phenomenon. Major extinctions in terrestrial vertebrates and large amphibians. Earliest examples of armoured dinosaurs. |
| 195 Ma | First pterosaurs with specialized feeding (Dorygnathus). First sauropod dinosaurs. Diversification in small, ornithischian dinosaurs: heterodontosaurids, fabrosaurids, and scelidosaurids. |
| 190 Ma | Pliosauroids appear in the fossil record. First lepidopteran insects (Archaeolepis), hermit crabs, modern starfish, irregular echinoids, corbulid bivalves, and tubulipore bryozoans. Extensive development of sponge reefs. |
| 176 Ma | First Stegosaurian dinosaurs. |
| 170 Ma | Earliest salamanders, newts, cryptoclidids, elasmosaurid plesiosaurs, and cladotherian mammals. Sauropod dinosaurs diversify. |
| 168 Ma | First lizards. |
| 165 Ma | First rays and glycymeridid bivalves. First vampire squids. |
| 163 Ma | Pterodactyloid pterosaurs first appear. |
| 161 Ma | Ceratopsian dinosaurs appear in the fossil record (Yinlong) and the oldest known eutherian mammal: Juramaia. |
| 160 Ma | Multituberculate mammals (genus Rugosodon) appear in eastern China. |
| 155 Ma | First blood-sucking insects (ceratopogonids), rudist bivalves, and cheilostome bryozoans. Archaeopteryx, a possible ancestor to the birds, appears in the fossil record, along with triconodontid and symmetrodont mammals. Diversity in stegosaurian and theropod dinosaurs. |
| 131 Ma | First pine trees. |
| 140 Ma | Orb-weaver spiders appear. |
| 135 Ma | Rise of the angiosperms. Some of these flowering plants bear structures that attract insects and other animals to spread pollen; other angiosperms are pollinated by wind or water. This innovation causes a major burst of animal coevolution. First freshwater pelomedusid turtles. Earliest krill. |
| 120 Ma | Oldest fossils of heterokonts, including both marine diatoms and silicoflagellates. |
| 115 Ma | First monotreme mammals. |
| 114 Ma | Earliest bees. |
| 112 Ma | Xiphactinus, a large predatory fish, appears in the fossil record. |
| 110 Ma | First hesperornithes, toothed diving birds. Earliest limopsid, verticordiid, and thyasirid bivalves. |
| 100 Ma | First ants. |
| 100–95 Ma | Spinosaurus appears in the fossil record. |
| 95 Ma | First crocodilians evolve. |
| 90 Ma | Extinction of ichthyosaurs. Earliest snakes and nuculanid bivalves. Large diversification in angiosperms: magnoliids, rosids, hamamelidids, monocots, and ginger. Earliest examples of ticks. Probable origins of placental mammals (earliest undisputed fossil evidence is 66 Ma). |
| 86–76 Ma | Diversification of therian mammals. |
| 70 Ma | Multituberculate mammals increase in diversity. First yoldiid bivalves. First possible ungulates (Protungulatum). |
| 68–66 Ma | Tyrannosaurus, the largest terrestrial predator of western North America, appears in the fossil record. First species of Triceratops. |
Cenozoic Era
Main article: Cenozoic
|
|
This section needs additional citations for verification. Please help improve this article by adding citations to reliable sources in this section. Unsourced material may be challenged and removed.
Find sources: "Timeline of the evolutionary history of life" – news · newspapers · books · scholar · JSTOR (September 2022) (Learn how and when to remove this message) |




.jpg/250px-Diprotodon_optatum_(2).jpg)

.jpg/250px-PantheraLeoAtrox1_(retouched).jpg)
| Date | Event |
|---|---|
| 66 Ma | The Cretaceous–Paleogene extinction event eradicates about half of all animal species, including mosasaurs, pterosaurs, plesiosaurs, ammonites, belemnites, rudist and inoceramid bivalves, most planktic foraminifers, and all of the dinosaurs excluding the birds. |
| 66 Ma | Rapid dominance of conifers and ginkgos in high latitudes, along with mammals becoming the dominant species. First psammobiid bivalves. Earliest rodents. Rapid diversification in ants. |
| 63 Ma | Evolution of the creodonts, an important group of meat-eating (carnivorous) mammals. |
| 62 Ma | Evolution of the first penguins. |
| 60 Ma | Diversification of large, flightless birds. Earliest true primates,[who?] along with the first semelid bivalves, edentate, carnivoran and lipotyphlan mammals, and owls. The ancestors of the carnivorous mammals (miacids) were alive.[citation needed] |
| 59 Ma | Earliest sailfish appear. |
| 56 Ma | Gastornis, a large flightless bird, appears in the fossil record. |
| 55 Ma | Modern bird groups diversify (first song birds, parrots, loons, swifts, woodpeckers), first whale (Himalayacetus), earliest lagomorphs, armadillos, appearance of sirenian, proboscidean mammals in the fossil record. Flowering plants continue to diversify. The ancestor (according to theory) of the species in the genus Carcharodon, the early mako shark Isurus hastalis, is alive. Ungulates split into artiodactyla and perissodactyla, with some members of the former returning to the sea. |
| 52 Ma | First bats appear (Onychonycteris). |
| 50 Ma | Peak diversity of dinoflagellates and nannofossils, increase in diversity of anomalodesmatan and heteroconch bivalves, brontotheres, tapirs, rhinoceroses, and camels appear in the fossil record, diversification of primates. |
| 40 Ma | Modern-type butterflies and moths appear. Extinction of Gastornis. Basilosaurus, one of the first of the giant whales, appeared in the fossil record. |
| 38 Ma | Earliest bears. |
| 37 Ma | First nimravid ("false saber-toothed cats") carnivores — these species are unrelated to modern-type felines. First alligators and ruminants. |
| 35 Ma | Grasses diversify from among the monocot angiosperms; grasslands begin to expand. Slight increase in diversity of cold-tolerant ostracods and foraminifers, along with major extinctions of gastropods, reptiles, amphibians, and multituberculate mammals. Many modern mammal groups begin to appear: first glyptodonts, ground sloths, canids, peccaries, and the first eagles and hawks. Diversity in toothed and baleen whales. |
| 33 Ma | Evolution of the thylacinid marsupials (Badjcinus). |
| 30 Ma | First balanids and eucalypts, extinction of embrithopod and brontothere mammals, earliest pigs and cats. |
| 28 Ma | Paraceratherium appears in the fossil record, the largest terrestrial mammal that ever lived. First pelicans. |
| 25 Ma | Pelagornis sandersi appears in the fossil record, the largest flying bird that ever lived. |
| 25 Ma | First deer. |
| 24 Ma | First pinnipeds. |
| 23 Ma | Earliest ostriches, trees representative of most major groups of oaks have appeared by now. |
| 20 Ma | First giraffes, hyenas, and giant anteaters, increase in bird diversity. |
| 17 Ma | First birds of the genus Corvus (crows). |
| 15 Ma | Genus Mammut appears in the fossil record, first bovids and kangaroos, diversity in Australian megafauna. |
| 10 Ma | Grasslands and savannas are established, diversity in insects, especially ants and termites, horses increase in body size and develop high-crowned teeth, major diversification in grassland mammals and snakes. |
| 9.5 Ma [dubious – discuss] |
Great American Interchange, where various land and freshwater faunas migrated between North and South America. Armadillos, opossums, hummingbirds Phorusrhacids, Ground Sloths, Glyptodonts, and Meridiungulates traveled to North America, while horses, tapirs, saber-toothed cats, jaguars, bears, coaties, ferrets, otters, skunks and deer entered South America. |
| 9 Ma | First platypuses. |
| 6.5 Ma | First hominins (Sahelanthropus). |
| 6 Ma | Australopithecines diversify (Orrorin, Ardipithecus). |
| 5 Ma | First tree sloths and hippopotami, diversification of grazing herbivores like zebras and elephants, large carnivorous mammals like lions and the genus Canis, burrowing rodents, kangaroos, birds, and small carnivores, vultures increase in size, decrease in the number of perissodactyl mammals. Extinction of nimravid carnivores. First leopard seals. |
| 4.8 Ma | Mammoths appear in the fossil record. |
| 4.5 Ma | Marine iguanas diverge from land iguanas. |
| 4 Ma | Australopithecus evolves. Stupendemys appears in the fossil record as the largest freshwater turtle, first modern elephants, giraffes, zebras, lions, rhinoceros and gazelles appear in the fossil record |
| 3.6 Ma | Blue whales grow to modern size. |
| 3 Ma | Earliest swordfish. |
| 2.7 Ma | Paranthropus evolves. |
| 2.5 Ma | Earliest species of Arctodus and Smilodon evolve. |
| 2 Ma | First members of genus Homo, Homo Habilis, appear in the fossil record. Diversification of conifers in high latitudes. The eventual ancestor of cattle, aurochs (Bos primigenus), evolves in India. |
| 1.7 Ma | Australopithecines go extinct. |
| 1.2 Ma | Evolution of Homo antecessor. The last members of Paranthropus die out. |
| 1.0 Ma | First coyotes. |
| 810 ka | First wolves |
| 600 ka | Evolution of Homo heidelbergensis. |
| 400 ka | First polar bears. |
| 350 ka | Evolution of Neanderthals. |
| 300 ka | Gigantopithecus, a giant relative of the orangutan from Asia dies out. |
| 250 ka | Anatomically modern humans appear in Africa. Around 50 ka they start colonising the other continents, replacing Neanderthals in Europe and other hominins in Asia. |
| 70 ka | Genetic bottleneck in humans (Toba catastrophe theory). |
| 40 ka | Last giant monitor lizards (Varanus priscus) die out. |
| 35–25 ka | Extinction of Neanderthals. Domestication of dogs. |
| 15 ka | Last woolly rhinoceros (Coelodonta antiquitatis) are believed to have gone extinct. |
| 11 ka | Short-faced bears vanish from North America, with the last giant ground sloths dying out. All Equidae become extinct in North America. Domestication of various ungulates. |
| 10 ka | Holocene epoch starts after the Last Glacial Maximum. Last mainland species of woolly mammoth (Mammuthus primigenus) die out, as does the last Smilodon species. |
| 8 ka | The giant lemur dies out. |
Plant evolution is an aspect of the study of biological evolution, predominantly involving evolution of plants suited to live on land, greening of various land masses by the filling of their niches with land plants, and diversification of groups of land plants.
Earliest plants
In the strictest sense, the name plant refers to those land plants that form the clade Embryophyta, comprising the bryophytes and vascular plants. However, the clade Viridiplantae or green plants includes some other groups of photosynthetic eukaryotes, including green algae. It is widely believed that land plants evolved from a group of charophytes, most likely simple single-celled terrestrial algae similar to extant Klebsormidiophyceae.
Chloroplasts in plants evolved from an endosymbiotic relationship between a cyanobacterium, a photosynthesising prokaryote and a non-photosynthetic eukaryotic organism, producing a lineage of photosynthesizing eukaryotic organisms in marine and freshwater environments. These earliest photosynthesizing single-celled autotrophs evolved into multicellular organisms such as the Charophyta, a group of freshwater green algae.
Fossil evidence of plants begins around 3000 Ma with indirect evidence of oxygen-producing photosynthesis in the geological record, in the form of chemical and isotopic signatures in rocks and fossil evidence of colonies of cyanobacteria, photosynthesizing prokaryotic organisms. Cyanobacteria use water as a reducing agent, producing atmospheric oxygen as a byproduct, and they thereby profoundly changed the early reducing atmosphere of the earth to one in which modern aerobic organisms eventually evolved. This oxygen liberated by cyanobacteria then oxidized dissolved iron in the oceans, the iron precipitated out of the sea water, and fell to the ocean floor to form sedimentary layers of oxidized iron called Banded Iron Formations (BIFs). These BIFs are part of the geological record of evidence for the evolutionary history of plants by identifying when photosynthesis originated. This also provides deep time constraints upon when enough oxygen could have been available in the atmosphere to produce the ultraviolet blocking stratospheric ozone layer. The oxygen concentration in the ancient atmosphere subsequently rose, acting as a poison for anaerobic organisms, and resulting in a highly oxidizing atmosphere, and opening up niches on land for occupation by aerobic organisms.
Fossil evidence for cyanobacteria also comes from the presence of stromatolites in the fossil record deep into the Precambrian. Stromatolites are layered structures formed by the trapping, binding, and cementation of sedimentary grains by microbial biofilms, such as those produced by cyanobacteria. The direct evidence for cyanobacteria is less certain than the evidence for their presence as primary producers of atmospheric oxygen. Modern stromatolites containing cyanobacteria can be found on the west coast of Australia and other areas in saline lagoons and in freshwater.
|
Key innovations in
early plant evolution |
||||||||||||||||||||||
|
−450 —
–
−440 —
–
−430 —
–
−420 —
–
−410 —
–
−400 —
–
−390 —
–
−380 —
–
−370 —
–
−360 —
|
O
S
D
L
Ll
W
Lu
Pridoli
E
M
L
Sandbian
Katian
Hirnantian
Rhuddanian
Aeronian
Telychian
Sheinwoodian
Homerian
Gorstian
Ludfordian
Lochkovian
Pragian
Emsian
Eifelian
Givetian
Frasnian
Famennian
|
|
||||||||||||||||||||
|
Axis scale: millions of years ago.
|
||||||||||||||||||||||
Paleozoic flora
Cambrian flora
Early plants were small, unicellular or filamentous, with simple branching. The identification of plant fossils in Cambrian strata is an uncertain area in the evolutionary history of plants because of the small and soft-bodied nature of these plants. It is also difficult in a fossil of this age to distinguish among various similar appearing groups with simple branching patterns, and not all of these groups are plants. One exception to the uncertainty of fossils from this age is the group of calcareous green algae, Dasycladales found in the fossil record since the middle Cambrian. These algae do not belong to the lineage that is ancestral to the land plants. Other major groups of green algae had been established by this time, but there were no land plants with vascular tissues until the mid-Silurian.
Ordovician flora
The evidence of plant evolution changes dramatically in the Ordovician with the first extensive appearance of embryophyte spores in the fossil record. The earliest terrestrial plants lived during the Middle Ordovician around 470 million years ago, based on their fossils found in the form of monads and spores, with resistant polymers in their outer walls, from Turkey, Saudi Arabia and Argentina. Individual trilete spores resembling those of modern cryptogamic plants and vascular plants first appeared in the fossil record from the Late Ordovician. These plants probably resembled liverworts, and did not have any conducting tissues. They were able to reproduce with spores, important dispersal units that have hard protective outer coatings which not only allowed their preservation in the fossil record, but also protected them from the UV light, desiccating environment and possible microorganism attack.
Silurian flora

The first fossil records of vascular plants, that is, land plants with vascular tissues, appeared in the Silurian period. The earliest known representatives of this group (mostly from the northern hemisphere) are placed in the genus Cooksonia. They had very simple branching patterns, with the branches terminated by flattened sporangia. By the end of the Silurian much more complex vascular plants, the zosterophylls, had diversified and primitive lycopods, such as Baragwanathia (originally discovered in Silurian deposits in Victoria, Australia), had become widespread.
Devonian flora
By the Devonian Period, the colonization of the land by plants was well underway. The bacterial and algal mats were joined early in the period by primitive plants that created the first recognizable soils and harbored some arthropods like mites, scorpions and myriapods. Early Devonian plants did not have roots or leaves like the plants most common today, and many had no vascular tissue at all. They probably relied on arbuscular mycorrhizal symbioses with fungi to provide them with water and mineral nutrients such as phosphorus. They probably spread by a combination of vegetative reproduction forming clonal colonies, and sexual reproduction via spores and did not grow much more than a few centimeters tall.
By the Late Devonian, forests of large, primitive plants existed: lycophytes, sphenophytes, ferns, and progymnosperms had evolved. Most of these plants have true roots and leaves, and many were quite tall. The tree-like Archaeopteris, ancestral to the gymnosperms, and the giant cladoxylopsid trees had true wood. These are the oldest known trees of the world's first forests. Prototaxites was the fruiting body of an enormous fungus that stood more than 8 meters tall. By the end of the Devonian, the first seed-forming plants had appeared. This rapid appearance of so many plant groups and growth forms has been called the "Devonian Explosion". The primitive arthropods co-evolved with this diversified terrestrial vegetation structure. The evolving co-dependence of insects and seed-plants that characterizes a recognizably modern world had its genesis in the late Devonian. The development of soils and plant root systems probably led to changes in the speed and pattern of erosion and sediment deposition.
The 'greening' of the continents acted as a carbon dioxide sink, and atmospheric concentrations of this greenhouse gas may have dropped. This may have cooled the climate and led to a massive extinction event. see Late Devonian extinction.
Also in the Devonian, both vertebrates and arthropods were solidly established on the land.
Carboniferous flora


Early Carboniferous land plants were very similar to those of the preceding Latest Devonian, but new groups also appeared at this time.
The main Early Carboniferous plants were the Equisetales (Horse-tails), Sphenophyllales (scrambling plants), Lycopodiales (Club mosses), Lepidodendrales (arborescent clubmosses or scale trees), Filicales (Ferns), Medullosales (previously included in the "seed ferns", an artificial assemblage of a number of early gymnosperm groups) and the Cordaitales. These continued to dominate throughout the period, but during late Carboniferous, several other groups, Cycadophyta (cycads), the Callistophytales (another group of "seed ferns"), and the Voltziales (related to and sometimes included under the conifers), appeared.
The Carboniferous lycophytes of the order Lepidodendrales, which were cousins (but not ancestors) of the tiny club-mosses of today, were huge trees with trunks 30 meters high and up to 1.5 meters in diameter. These included Lepidodendron (with its fruit cone called Lepidostrobus), Halonia, Lepidophloios and Sigillaria. The roots of several of these forms are known as Stigmaria.
The fronds of some Carboniferous ferns are almost identical with those of living species. Probably many species were epiphytic. Fossil ferns include Pecopteris and the tree ferns Megaphyton and Caulopteris. Seed ferns or Pteridospermatophyta include Cyclopteris, Neuropteris, Alethopteris, and Sphenopteris.
The Equisetales included the common giant form Calamites, with a trunk diameter of 30 to 60 cm and a height of up to 20 meters. Sphenophyllum was a slender climbing plant with whorls of leaves, which was probably related both to the calamites and the modern horsetails.
Cordaites, a tall plant (6 to over 30 meters) with strap-like leaves, was related to the cycads and conifers; the catkin-like inflorescence, which bore yew-like berries, is called Cardiocarpus. These plants were thought to live in swamps and mangroves. True coniferous trees (Walchia, of the order Voltziales) appear later in the Carboniferous, and preferred higher drier ground.
Permian flora
The Permian began with the Carboniferous flora still flourishing. About the middle of the Permian there was a major transition in vegetation. The swamp-loving lycopod trees of the Carboniferous, such as Lepidodendron and Sigillaria, were replaced by the more advanced conifers, which were better adapted to the changing climatic conditions. Lycopods and swamp forests still dominated the South China continent because it was an isolated continent and it sat near or at the equator. The Permian saw the radiation of many important conifer groups, including the ancestors of many present-day families. The ginkgos and cycads also appeared during this period. Rich forests were present in many areas, with a diverse mix of plant groups. The gigantopterids thrived during this time; some of these may have been part of the ancestral flowering plant lineage, though flowers evolved only considerably later.
Mesozoic flora
Triassic flora
On land, the holdover plants included the lycophytes, the dominant cycads, Ginkgophyta (represented in modern times by Ginkgo biloba) and glossopterids. The spermatophytes, or seed plants came to dominate the terrestrial flora: in the northern hemisphere, conifers flourished. Dicroidium (a seed fern) was the dominant southern hemisphere tree during the Early Triassic period.
Jurassic flora
The arid, continental conditions characteristic of the Triassic steadily eased during the Jurassic period, especially at higher latitudes; the warm, humid climate allowed lush jungles to cover much of the landscape. Conifers dominated the flora, as during the Triassic; they were the most diverse group and constituted the majority of large trees. Extant conifer families that flourished during the Jurassic included the Araucariaceae, Cephalotaxaceae, Pinaceae, Podocarpaceae, Taxaceae and Taxodiaceae. The extinct Mesozoic conifer family Cheirolepidiaceae dominated low latitude vegetation, as did the shrubby Bennettitales. Cycads were also common, as were ginkgos and tree ferns in the forest. Smaller ferns were probably the dominant undergrowth. Caytoniaceous seed ferns were another group of important plants during this time and are thought to have been shrub to small-tree sized. Ginkgo-like plants were particularly common in the mid- to high northern latitudes. In the Southern Hemisphere, podocarps were especially successful, while Ginkgos and Czekanowskiales were rare.
Cretaceous flora
Flowering plants, also known as angiosperms, spread during this period, although they did not become predominant until near the end of the period (Campanian age). Their evolution was aided by the appearance of bees; in fact angiosperms and insects are a good example of coevolution. The first representatives of many modern trees, including figs, planes and magnolias, appeared in the Cretaceous. At the same time, some earlier Mesozoic gymnosperms, like Conifers continued to thrive, although other taxa like Bennettitales died out before the end of the period.